Роль генотипа и условий внешней среды в формировании фенотипа. Роль генотипа и условий среды в формировании поведенческих признаков у животных
Роль генотипа и условий внешней среды в формировании фенотипа
Роль генотипа и условий внешней среды в формировании фенотипа
Все многообразие всего живого и его постоянное совершенствование были бы невозможны без изменчивости. Это связано с тем, что генотип последовательно реализуется в фенотип в ходе индивидуального развития организма и в определенных условиях среды обитания, факторы которой (колебания освещенности, температуры, влажности, условий питания, взаимоотношений с другими организмами и др.) часто оказывают определяющее значение на проявление и развитие того или иного признака и свойства. Поэтому организмы, имеющие одинаковые генотипы, могут заметно отличаться друг от друга по фенотипу. Приведем несколько примеров.
Если растения примулы, которые в обычных условиях имели красные цветки, перенести в оранжерею и содержать их там при температуре 30—35°С и высокой влажности, то через некоторое время все цветки у них оказываются белыми. Если эти же растения вновь поместить в обычные (комнатные) условия, то распустившиеся цветки будут красными.
Другим классическим примером, иллюстрирующим влияние внешней среды на проявление качественных признаков, служит изменение окраски шерсти у гималайского кролика. Обычно при 20°С у этого кролика шерсть белая, за исключением черных ушей, лап, хвоста и мордочки. При 30°С такие кролики вырастают полностью белыми. Если же у гималайского кролика сбрить шерсть на боку или спине и содержать его при температуре воздуха ниже +2°С, то вместо белой шерсти вырастет черная. Но если сбрить шерсть на ухе, то в обычных условиях там снова вырастет черная шерсть.
Эти наблюдения объясняют, почему гималайские кролики рождаются белыми, без участков черной шерсти: их эмбриональное развитие происходит в условиях высокой температуры.
Но даже в том случае, когда проявление и развитие того или иного признака происходит, степень его выраженности бывает
разной в зависимости от условий внешней среды: при одних она усиливается, при других ослабляется. Пределы этих изменений определены возможностями, заложенными в генотипе.
Пределы модификационной изменчивости признака, ограниченные действием генотипа, называют его норной реакции.
Для разных признаков и свойств организмов границы, определяемые нормой реакции, неодинаковы. Наибольшей пластичностью и изменчивостью характеризуются количественные признаки: семенная продуктивность злаков, величина удоя у крупного рогатого скота, масса животных, число и размеры листьев и колосков у растений и т. д. В то же время качественные признаки (масть животных, окраска семян, цветков и плодов, остистость и опушенность колоса у растений) мало зависят от условий среды.
Так, у крупного рогатого скота удой во многом определяется условиями его содержания. При подборе кормов нужного качества и количества ценная порода скота может дать 5—6 тыс. кг молока в год, а в случае ухудшения условий содержания продуктивность снижается до 2,5 тыс. кг и даже ниже. Что касается масти животных, то при самых разных условиях она почти не изменяется.
Подобную закономерность можно наблюдать и у растений. Например, растения элитного сорта картофеля, выращенные в условиях высокой культуры агротехники, дают урожай клубней 500—600 ц/га и более. Ухудшение условий (тяжелые глинистые почвы, низкая культура земледелия и ухода за растениями) может привести к тому, что урожайность прекрасного сорта картофеля снизится до 100—150 ц/га и ниже. Правда, окраска клубней и их форма при этом изменяются в очень малой степени.
Рассмотренные примеры свидетельствуют о том, что количественные признаки обладают очень широкой нормой реакции, а качественные — более узкой.
Таким образом, все признаки и свойства каждого сорта растений и каждой породы животных могут изменяться только в пределах нормы реакции. Поэтому попытки добиться высоких урожаев, резко повысить продуктивность сельскохозяйственных животных только за счет повышения уровня культуры земледелия, улучшения условий кормления и содержания животных несостоятельны. В первую очередь необходимо получить новые сорта растений и породы животных улучшенного генотипа, отзывчивые на хорошие условия возделывания и выращивания.
По механизму возникновения и характеру изменений признаков различают два типа изменчивости — наследственную и ненаследственную.
Под наследственной изменчивостью понимают способность к изменениям самого генетического материала, а под ненаследственной (модификационной, или фенотипической) — способность организмов реагировать на условия окружающей среды и изменяться в пределах нормы реакции, определяемой генотипом.
sbio.info
Какова роль генотипа и условий внешней среды в формировании фенотипа?
Какова роль генотипа и условий внешней среды в формировании фенотипа?
Генотип — совокупность генов одного организма, но генотип не механическая сумма всех генов. Возможность проявления Гена и форма его проявления зависит от среды. В понятие среды входят не только условия, окружающие клетку, но и другие гены. Гены взаимодействуют друг с другом и, оказавшись в одном генотипе, могут сильно влиять на проявление действия соседних генов, т.е. для каждого гена существует генотипическая среда. Поэтому М.Е. Лобашев определил генотип как систему взаимодействующих генов. Гены проявляются в виде признаков. Возможность проявления зависит от условий окружающей среды. Совокупность всех признаков организма — фенотип. Сюда относят не только внешние видимые признаки (цвет волос, глаз), но и биохимические (форма молекулы белка или фермента), гистологические (форма и размеры клеток, строение тканей и органов).
Фенотип формируется под влиянием генотипа и условий внешней среды. При этом и качественные, и количественные признаки подвержены влиянию внешней среды. У примулы окраска цветков определяется аллельной парой Rr. Гомозиготные растения RR обычно имеют красные цветки, но если в момент формирования бутонов растение перенести из обычных комнатных условий в теплую влажную оранжерею с температурой +30° — +35°С, то появятся белые цветки. Возвращение в комнатные условия не изменяет их белой окраски, но вновь распустившиеся цветки будут красными. Понятно, что в этом случае изменился признак, а не ген.
Развитие количественных признаков очень сильно зависит от влияния условий среды. Масса тела крупного рогатого скота — типичный количественный признак. Установлено, что генотип оказывает важное влияние на формирование признака. Именно благодаря различиям в генотипе породы крупного рогатого скота резко отличаются по среднему значению, например, массы одного животного. Однако условия среды, например количество и качество корма, играют не менее важную роль в формировании этого признака.
Какова роль генотипа и условий внешней среды в формировании фенотипа?
Оцените пожалуйста этот постНа этой странице искали :
- каковы условия проявления генотипа во внешнем облике организма
- роль генотипа и условий среды в формировании поведенческих признаков у животных
- роль генотипа и среды в формировании фенотипа
- Роль генотипа и среды в формировании организма ребенка
- какова роль генотипа и условий среды в формировании фенотипа
Сохрани к себе на стену!
vsesochineniya.ru
Роль генотипа и условий внешней среды в формировании фенотипа
Все многообразие всего живого и его постоянное совершенствование были бы невозможны без изменчивости. Это связано с тем, что генотип последовательно реализуется в фенотип в ходе индивидуального развития организма и в определенных условиях среды обитания, факторы которой (колебания освещенности, температуры, влажности, условий питания, взаимоотношений с другими организмами и др.) часто оказывают определяющее значение на проявление и развитие того или иного признака и свойства. Поэтому организмы, имеющие одинаковые генотипы, могут заметно отличаться друг от друга по фенотипу. Приведем несколько примеров.
Если растения примулы, которые в обычных условиях имели красные цветки, перенести в оранжерею и содержать их там при температуре 30—35°С и высокой влажности, то через некоторое время все цветки у них оказываются белыми. Если эти же растения вновь поместить в обычные (комнатные) условия, то распустившиеся цветки будут красными.
Другим классическим примером, иллюстрирующим влияние внешней среды на проявление качественных признаков, служит изменение окраски шерсти у гималайского кролика. Обычно при 20°С у этого кролика шерсть белая, за исключением черных ушей, лап, хвоста и мордочки. При 30°С такие кролики вырастают полностью белыми. Если же у гималайского кролика сбрить шерсть на боку или спине и содержать его при температуре воздуха ниже +2°С, то вместо белой шерсти вырастет черная. Но если сбрить шерсть на ухе, то в обычных условиях там снова вырастет черная шерсть.
Эти наблюдения объясняют, почему гималайские кролики рождаются белыми, без участков черной шерсти: их эмбриональное развитие происходит в условиях высокой температуры.
Известно, что все признаки и свойства организма наследственно детерминированы, однако организмы наследуют не сами признаки и свойства, а лишь возможность их развития. Для проявления и развития признака необходимы соответствующие условия внешней среды.
Но даже в том случае, когда проявление и развитие того или иного признака происходит, степень его выраженности бывает
разной в зависимости от условий внешней среды: при одних она усиливается, при других ослабляется. Пределы этих изменений определены возможностями, заложенными в генотипе.
Пределы модификационной изменчивости признака, ограниченные действием генотипа, называют его норной реакции.
Для разных признаков и свойств организмов границы, определяемые нормой реакции, неодинаковы. Наибольшей пластичностью и изменчивостью характеризуются количественные признаки: семенная продуктивность злаков, величина удоя у крупного рогатого скота, масса животных, число и размеры листьев и колосков у растений и т. д. В то же время качественные признаки (масть животных, окраска семян, цветков и плодов, остистость и опушенность колоса у растений) мало зависят от условий среды.
Так, у крупного рогатого скота удой во многом определяется условиями его содержания. При подборе кормов нужного качества и количества ценная порода скота может дать 5—6 тыс. кг молока в год, а в случае ухудшения условий содержания продуктивность снижается до 2,5 тыс. кг и даже ниже. Что касается масти животных, то при самых разных условиях она почти не изменяется.
Подобную закономерность можно наблюдать и у растений. Например, растения элитного сорта картофеля, выращенные в условиях высокой культуры агротехники, дают урожай клубней 500—600 ц/га и более. Ухудшение условий (тяжелые глинистые почвы, низкая культура земледелия и ухода за растениями) может привести к тому, что урожайность прекрасного сорта картофеля снизится до 100—150 ц/га и ниже. Правда, окраска клубней и их форма при этом изменяются в очень малой степени.
Рассмотренные примеры свидетельствуют о том, что количественные признаки обладают очень широкой нормой реакции, а качественные — более узкой.
Таким образом, все признаки и свойства каждого сорта растений и каждой породы животных могут изменяться только в пределах нормы реакции. Поэтому попытки добиться высоких урожаев, резко повысить продуктивность сельскохозяйственных животных только за счет повышения уровня культуры земледелия, улучшения условий кормления и содержания животных несостоятельны. В первую очередь необходимо получить новые сорта растений и породы животных улучшенного генотипа, отзывчивые на хорошие условия возделывания и выращивания.
По механизму возникновения и характеру изменений признаков различают два типа изменчивости — наследственную и ненаследственную.
Под наследственной изменчивостью понимают способность к изменениям самого генетического материала, а под ненаследственной (модификационной, или фенотипической) — способность организмов реагировать на условия окружающей среды и изменяться в пределах нормы реакции, определяемой генотипом.
mirznanii.com
Роль генотипа и внешней среды в проявлении признаков.
Прогресс в понимании причин и источников явлений изменчивости и наследственности у человека, включая и патологическую, в значительной мере обязан медицинской и клинической генетике. Главным фактором, определяющим развитие любого признака, является генотип. Генотип организма формируется в момент оплодотворения, но степень последующего формирования генетического потенциала в значительной степени зависит от воздействующих на организм внешних факторов. Примером свидетельства влияния условий внешней среды на развитие генотипа у человека являются результаты исследования близнецов: если развитие одного из близнецов проходило в благоприятных условиях по сравнению с другим близнецом, то его физическое и интеллектуальное развитие будет выше, чем у близнеца, воспитанного в неблагоприятных условиях. Это позволяет сделать вывод, что на фенотипическое проявление признака оказывают влияние, как наследственность, так и условия внешней среды. Кроме того, наблюдения позволяют определить непрерывную фенотипическую изменчивость как накопительный результат различных факторов среды, воздействующих на вариабельный генотип и степень наследуемости данного признака определяется прежде всего генотипом.
Организм человека, включающий большое количество разнообразных белков, функционирует как единая система, в которой развитие одних структур зависит от функции других и необходимости приспособления к различным меняющимся условиям внешней среды. Даже для некоторых заболеваний, развитие которых контролируется одним конкретным геном, удается изменить характер патологического процесса, изменяя условия среды.
Например, при наследственном моногенном заболевании галактоземии, организм ребенка не может нормально использовать галактозу, которую он получает в составе молочного сахара с материнским или коровьим молоком. Обычное вскармливание ребенка с галактоземией развивает слабоумие, поражение печени и глаз. При исключении галактозы из питания дети, несмотря на аномальную генетическую конституцию, могут нормально развиваться. Таким образом, фактор внешней среды, в данном случае исключение галактозы из питания, меняет фенотипическое действие гена.
Условия, в которых существует организм человека, могут модифицировать полигеннодетерминированные признаки. Например, рост ребенка контролируется целым рядом пар нормальных генов, регулирующих обмен гормонов, минералов, пищеварение и т.д. Но далее если изначально генетически определен высокий рост, а человек живет в плохих условиях (недостаток солнца, воздуха, неполноценное питание), то это приводит к низкорослости.
Отдельно только генотип или только факторы внешней среды не могут определить формирование фенотипических характеристик какого-либо признака. Что касается развития интеллекта, таланта и индивидуальности, то судя по имеющимся данным, они зависят как от наследственности, так и от среды, которая влияет на окончательную выраженность признака. Важной задачей генетики является уточнение роли наследственных и внешнесредовых факторов в формировании того или иного признака. Необходимо оценить степень обусловленности количественных характеристик организма генетическими различиями между особями или средовой изменчивостью. Для количественной оценки этих воздействий американский генетик Дж. Лаш ввел термин «наследуемость».
Наследуемость отражает вклад генетических факторов в фенотипическое проявление конкретного признака. Этот показатель может иметь значение от 0 до 1 (0—100%). Чем ниже уровень наследуемости, тем меньше роль генотипа в изменчивости данного признака. Если наследуемость приближается к 100%, то фенотипическая изменчивость признака почти полностью определяется наследственными факторами.
Дата добавления: 2015-10-02; просмотров: 976 | Нарушение авторских прав
Аллельные и неаллельные гены | Законы Менделя | Качественных характеристик | Типы наследования менделирующих признаков у человека | Генотип. Фенотип | Количественная и качественная специфика проявления генов в признаках | Хромосомная теория наследственности | Человека | Составление родословной | Генеалогический анализ родословной. |mybiblioteka.su - 2015-2018 год. (0.015 сек.)mybiblioteka.su
Наследование поведения животными
Полигенное наследование поведения
Многие поведенческие признаки контролируются большим числом генов и большим числом факторов окружающей среды. Для генетического анализа таких сложных ситуаций можно использовать разнообразные методы. Их эффективность в большой мере зависит от степени наследуемости рассматриваемого поведенческого признака. Однако некоторые аспекты поведения характеризуются высокой степенью наследуемости, с них мы и начнем обсуждение полигенного наследования.
Самцы сверчков привлекают самок с большого расстояния призывными сигналами. Звук извлекается при ритмичном раскрывании и складывании специализированных надкрылий, на которых находится специальный фрикционный механизм. При каждом складывании надкрылий возникает звук, тогда как раскрывание беззвучно. Эти призывные сигналы удивительно стереотипны у членов локальной популяции, но они резко различаются у разных видов. Различия имеют место в основном во временной организации звуковых пульсов. Межвидовые гибриды в природе не встречаются (Hill et al., 1972), однако их можно получить в лаборатории. Например, Леруа (Leroy, 1964) получил гибрид между австралийскими полевыми сверчками Teleogryllus commodus и Т. oceanicus. Бентли и Хой (Bentley, Hoy, 1972) обнаружили, что призывные сигналы гибридов первого поколения (Fj) заметно отличаются от призывных сигналов обоих этих видов. В частности, интервалы между сериями и между трелями гибридов имели значения, промежуточные между значениями соответствующих интервалов у родителей. На рис. 4.5 сравниваются осциллограммы призывных сигналов Т. commodus и их гибридов. По данным Бентли и Хоя, реципрокные гибриды отличаются друг от друга. Призывный сигнал гибрида, полученного от самки Т. oceanicus и самца Т. commodus, был похож на сигнал Т. oceanicus наличием отчетливо выраженного интервала между трелями. У гибрида, полученного от самки Т. commodus и самца Т. oceanicus, в звуковом сигнале отсутствует хорошо выраженный интервал между трелями, и он похож на сигнал Т. commodus. Эти различия лучше всего видны на гистограмме распределения межимпульсных интервалов (рис. 4.6.).
Эти результаты позволяют предположить, что наследование характера при зывного сигнала полигенно. Мы не располагаем данными о том, что какой-либо конкретный признак призывного сигнала является доминантным и призывные сигналы гибридов промежуточны по своим параметрам между призывными сигналами родителей.
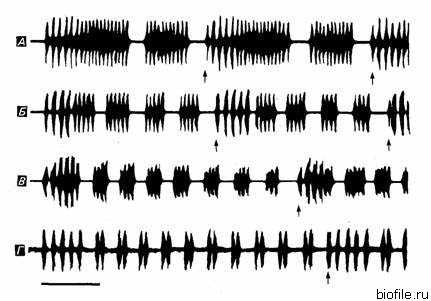
Рис. 4.5. Осциллограммы призывных сигналов Т. oceanicus (А.). Т. commodus (Г) и их гибридов: самка Т. oceanicus х самец Т. commodus (Б) и самка Т. commodus х самец Т. oceanicus (В) (стрелками отмечено начало фраз). (По Bentley, Hoy, 1972.)
Эта гипотеза подкрепляется тем фактом, что у гибридов от возвратного скрещивания также наблюдается промежуточное наследование (Bentley, 1971). Кроме того, существуют некоторые указания на то, что наследование этих признаков сцеплено с полом. Призывные сигналы гибридов от реципрокного скрещивания отличаются таким образом, что можно думать, что интервалы между трелями, свойственные Т. oceanicus, есть у одних гибридов, но отсутствуют у других. Обозначение пола у сверчков, ХО, означает, что Y-хромосома отсутствует. Самец получает Х-хромосому от матери, но не получает ничего, что соответствовало бы ей от отца. Таким образом, оказывается, что интервал между трелями у гибрида от самки Т. oceanicus и самца Т. commodus определяется Х-хромосомой матери Т. oceanicus. Этого признака нет ни у гибридов от самки Т. commodus и самца Т. oceanicus, ни у их матерей.
На призывные сигналы самцов различных видов реагируют преимущественно самки соответствующего вида. Интересно, однако, узнать, как гибридные самки реагируют на призывные сигналы своих родителей и своих братьев. Самки сверчков немые и реагируют на призывные сигналы самца тем, что ползут по направлению к источнику звука. Хилл и др. (Hill et al., 1972) показали, что самки Т. oceanicus и Т. commodus, имеющие возможность свободно перемещаться, могут отличать призывный сигнал своего вида от призывного сигнала других видов.
Чтобы выразить количественно предпочтение самки, Хой и Пол (Ноу, Poul, 1973) использовали Y-образный лабиринт. Они воспроизводили записанные призывные сигналы через громкоговоритель, помещенный справа и слева от лабиринта. Таким способом они имели возможность измерять относительную привлекательность любого призывного сигнала. Хой и Пол обнаружили, что самки Т. oceanicus и Т. commodus отличают видовой призывный сигнал; это же было обнаружено Хиллом и др. у свободно перемещающихся самок. Кроме того, эти авторы показали, что гибридные самки, являющиеся дочерьми самок Т. oceanicus и самцов Т. commodus, призывный сигнал своих братьев предпочитают типичному призывному сигналу любого из родителей. Как видно из рис. 4.6, призывный сигнал гибрида от скрещивания самца Т. oceanicus и самки Т. commodus легко отличить от призывного сигнала любого из родителей. (Обратите внимание, что он не так четко разделен на интервалы, как в случае с гибридом от другой комбинации.) Тот факт, что призывный сигнал гибрида предпочитается самкой, означает, что призывный сигнал самца и восприятие и распознавание его самкой каким-то образом связаны генетически.
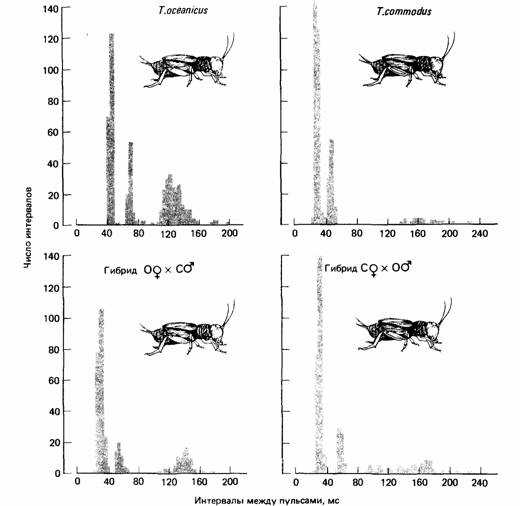
Рис. 4.6. Гистограммы частот интервалов между пульсами в призывных сигналах сверчков. Каждая гистограмма представляет собой анализ одного индивидуального сигнала. На гистограммах можно различить внутрисерийные интервалы, интервалы в пределах трели и интервалы между трелями. (По Bentley, Hoy, 1972.)
Хорошо известно, что различные породы собак характеризуются разными поведенческими признаками. В 1965 г. Скотт и Фуллер (Scott, Fuller, 1965) опубликовали некоторые результаты своих обширных исследований по генетической основе таких различий. В одном исследовании они сравнивали поведение коккер-спаниелей, басенджи и их гибридов. Спаниели с давних пор воспитывались как спортивные собаки, послушные и преданные своему хозяину. Родиной спаниелей считается Испания, где их использовали на охоте с ловчими птицами и при ловле птиц сетями. Собаку приучали прижиматься к земле, когда она обнаружит куропатку или другую дичь. Затем брошенной сетью накрывали и птицу, и собаку. После изобретения огнестрельного оружия сеть больше не использовали и спаниелей научили останавливаться и указывать, где они обнаружили птиц. Были выведены спрингер-спаниели, которые прыгали и вспугивали птиц. Первоначальная привычка затаиваться осталась только у коккер-спаниелей.
Басенджи широко распространены в Африке; пигмеи и некоторые другие африканские племена используют их на охоте. Это многоцелевые охотничьи собаки, довольно осторожные. Басенджи молчаливы и почти никогда не лают, хотя порой они долго и протяжно воют.
Скотт и Фуллер (Scott, Fuller, 1965) осуществили обширную программу скрещивания этих двух пород (рис. 4.10), включая реципрокные скрещивания и различные типы возвратного скрещивания. В табл. 4.1 суммированы признаки этих двух пород и указаны наиболее вероятные способы наследования, основанные на результатах экспериментов. Щенки басенджи обычно дикие в сравнении с ручными и ласковыми щенками спаниелей. Они избегают рук и сопротивляются, если их пытаются удержать. Проверка на приручение показала, что поведение щенков F1 сходно с поведением родителей и определяется, таким образом, доминантными генами. Результаты экспериментов по возвратному скрещиванию свидетельствуют о том, что единичный доминантный ген контролирует дикость, обычно свойственную басенджи. Способность к послушанию у спаниелей контролируется единичным рецессивным геном. Возьмем другой пример. У самок басенджи течка бывает обычно раз в год, во время осеннего равноденствия. У спаниелей, как и у большинства европейских пород, это происходит каждые шесть месяцев в любое время года. Эстральный цикл у басенджи, очевидно, контролируется единичным рецессивным геном. Учитывая все это, Скотт и Фуллер пришли к выводу, что исследуемые ими поведенческие признаки контролируются одним или двумя генами. Эта ситуация оказывается промежуточной между простым менделевским наследованием и полигенным наследованием.
Таблица 4.1.
|
Признаки |
Бассенджи |
Коккер-спаниели |
Наиболее вероятный способ наследования |
|
Избегание и голосовые реакции при взятии на руки |
Высокое |
Низкое |
Один доминантный ген, определяющий дикость |
|
Сопротивление ограничению движений |
Высокое |
Низкое |
Один ген без доминирования |
|
Агрессивность во время игры в возрасте 13-15 нед |
Высокая |
Низкая |
Два гена без доминирования |
|
Порог стимуляции |
Высокий |
Низкий |
Два доминантных гена, определяющие низкий порог |
|
Тенденция лаять малое число раз |
Высокая |
Низкая |
Один ген без доминирования |
|
Половое поведение (время течки) |
1 раз в год |
1 раз в полгода |
Тип басенджи определяется одним рецессивным геном |
|
Тенденция сохранять спокойствие при взвешивании |
Низкая |
Высокая |
Два рецессивных гена, определяющих сильную тенденцию. |
Может показаться удивительным, что такие сложные черты поведения контролируются таким малым числом генов. Однако эти две породы собак были изолированы в течение долгого времени и подвергались интенсивному искусственному отбору. Это, по-видимому, сформировало генотип, который гомозиготен по отдельным признакам, так что имеется лишь небольшое расщепление внутри пород. Генетика поведения собак особенно интересна, так как способность научиться конкретному типу поведения у них часто является наследственной.


Рис. 4.10. Родительские пары коккер-спаниелей (вверху) и басенджи (внизу), использованные в генетических исследованиях Скотта и Фуллера. (Skott, Fuller, 1965; фотография J. P. Scott.)
Наследование поведенческих признаков
«Вопрос о том, что важнее в формировании физических черт человека и его личности - природа или воспитание, генотип или окружающая среда, - попросту ошибочен и может ввести в заблуждение. Генотип и окружающая среда важны в равной степени, так как и то и другое необходимо... Вопрос о роли генотипа и окружающей среды в развитии человека следует поставить так: до какой степени различия, наблюдаемые между людьми, определяются различиями их генотипов и различиями условий, в которых люди рождаются, растут и воспитываются? (Dobzhansky, 1964).»
Большая часть работ Фрэнсиса Гальтона основана на представлении о том, что фенотипическое сходство между родственниками частично определяется сходством их генотипов. Однако Гальтон не был знаком с работами Менделя и считал, что наследственность носит слитный характер. Он думал, что материал наследственности непрерывен, поэтому признаки потомков промежуточны между признаками их родителей. На основе таких допущений Гальтон впервые разработал методику расчета корреляций между родственниками как средство оценки наследования тех или иных признаков. В 1918 г. Фишер опубликовал статью, в которой было показано, что возможно предсказать корреляции, ожидаемые между родственниками на основе теории Менделя. Работы Менделя стали известны общественности в 1900 г., однако они относились только к дискретным признакам. Фишер попытался применить менделевскую теорию для анализа непрерывных признаков, таких, как рост и масса. Интересно, что его выводы были сходны с выводами Гальтона. причем большая часть прогнозов в этих двух теориях была почти идентична.
Наследуемость впервые была определена как доля изменчивости, связанная с различиями в наследственности (Lush, 1940). Иными словами, когда мы анализируем конкретный признак в популяции животных, такой, например, как масса тела, мы определяем, какой вклад вносят в фенотипическое проявление этого признака генотипическая и средовая компоненты. Рассматривая родственные популяции в различных условиях, иногда можно оценить степень влияния на изменчивость особей факторов окружающей среды и генетических факторов.
Существуют разнообразные статистические методы оценки наследуемости поведения. Некоторые основаны на исследовании фенотипической изменчивости у генотипически однородных особей. Эта общая фенотипическая изменчивость генотипически идентичных особей сравнивается с общей фенотипической изменчивостью в природной генотипически разнообразной популяции. Такое сравнение дает соотношение, в котором противопоставляется генотипическая компонента фенотипического разнообразия средовой компоненте. При оценке с помощью этой статистической методики мы сталкиваемся с определенными трудностями, но она ясно показывает, насколько ошибочно относить один вид поведения к «наследственному», а другой - к «приобретенному». В случае если возникнет новая генная вариация или популяция попадает в другие условия среды, это соотношение может сильно измениться. Типы поведения, которые «генетически детерминированы» в одной среде (у монозиготных близнецов, воспитанных вместе, фенотипы оказываются сходными), могут быть подвержены и сильному влиянию средовых факторов, например климатических (у монозиготных близнецов, воспитанных порознь, фенотипы оказываются разными). Напротив, тип поведения, который «детерминирован средовыми факторами» (у всех генотипически различающихся особей одинаковый фенотип, если их воспитывают в одинаковых условиях), может, очевидно, в значительной степени подвергаться воздействию генетической компоненты в среде, в которой проявляется генотипическое разнообразие. Все эти взаимодействия позволяют лишь в строгих пределах определить степень наследуемости того или иного типа поведения, а также провести границу между врожденным и приобретенным поведением. Мы можем сказать, что данный тип поведения может наследоваться в 80% случаев при данном генотипе, в данной среде и в конкретной популяции, но мы не можем переносить эти выводы на другую популяцию или среду.
При оценке наследуемости ошибки могут проистекать из разных источников, наиболее важный из них - это взаимодействие генотипа и среды и корреляции генотипа и среды. Взаимодействие между генотипом и средой вводит изменчивость, которая не принимается во внимание при обычных расчетах наследуемости. Например, Трайон (Тгуоп, 1942) проводил селекцию крыс на неспособность ориентироваться в лабиринте («глупые» крысы) и на блестящее ориентирование в нем («умные» крысы). Этих крыс выводили и тестировали в обычных лабораторных условиях. А Купер и Зубек (Cooper, Zubek, 1958) выращивали одних крыс в обычных условиях, других - в обедненных, без укрытий и препятствий, а третьих - в условиях, обогащенных сложными лабиринтообразными сооружениями. При испытаниях в стандартном лабиринте эти две линии крыс, выращенные в обычных условиях, сильно различались по своим возможностям ориентироваться в лабиринте, как это обнаружили раньше и другие исследователи. Однако обе линии крыс, выращенных в обедненных условиях, одинаково плохо ориентировались в лабиринте, тогда как обе линии, выращенные в обогащенной среде, ориентировались одинаково хорошо. Хотя эти две линии отчетливо различаются генотипически, в фенотипе эта разница проявляется только у крыс, выращенных в определенных условиях среды.
Корреляция генотипа и среды - другой потенциальный источник ошибок при оценке наследуемости поведенческих признаков - может возрастать, когда особи избирают какую-то особую среду или у них развиваются определенные привычки для компенсации генотипических дефектов. Однако, несмотря на эти трудности, оценка наследуемости оказалась особенно полезной для разделения эффектов природы и воспитания.
Оценки наследуемости, основанные на сходстве между родственниками, часто использовались в генетическом анализе поведения человека. Однако здесь есть ряд осложняющих факторов. Во-первых, генетическое доминирование снижает корреляцию между родственниками. При генетическом доминировании вклады родителей в проявление признака неодинаковы, т. е. вводится еще один источник изменчивости. Это называется дисперсией доминирования. Однако дисперсия доминирования оказывает различное действие на корреляции родителей и потомков и корреляции между сибсами. Фактически разница между этими двумя коэффициентами корреляции должна точно составлять одну четверть дисперсии, обусловленной доминированием (Bodmer, Cavalli-Sforza, 1976).
Во-вторых, подбор пар у человека, вероятно, не является случайным в отношении некоторых признаков. Можно допустить, что в отношении определенной части признаков, которые нельзя наблюдать непосредственно, например уровней ферментов, подбор пар происходит случайным образом, однако этого нельзя сказать в отношении таких признаков, как рост и коэффициент умственного развития. Корреляции между мужем и женой по этим признакам обычно положительны, что указывает на неслучайный или выборочный подбор, поскольку люди обычно стремятся выбирать партнеров одинакового роста и умственного развития. Эти корреляции по росту составляют примерно +0,3, а по умственному развитию - примерно + 0,4. На эти корреляции могут влиять и социальные факторы, так как между социально-экономическим положением и данными признаками также существует положительная корреляция. Поэтому тенденция выбирать мужа или жену из той же социальной группы обычно приводит к положительной корреляции между мужем и женой. Ассортативные браки у человека могут влиять на увеличение частоты гомозигот.
В-третьих, при анализе наследуемости поведения человека необходимо правильно оценивать средовую дисперсию, поскольку в этом случае условия среды нельзя контролировать, как при исследованиях, проводимых на животных. Был предложен способ (Cavalli-Sforza, Bodmer, 1971) разделения средовой дисперсии на отдельные компоненты. Прежде всего следует различать внутрисемейную дисперсию. Она может быть обусловлена возрастными различиями, порядком рождения, различиями по полу и т.д. В семьях разных размеров эта дисперсия может также зависеть от уровня питания и наличия жилищной площади.
Далее существует дисперсия, обусловленная различиями между семьями одного социально-экономического слоя. Некоторую информацию о значении этого фактора можно почерпнуть из рассмотрения корреляции между приемными детьми и их приемными родителями, однако результаты могут быть тенденциозны из-за того, что родителей выбирают посреднические агентства.
Затем следует учитывать дисперсию между социально-экономическими слоями. Культурная преемственность может привести к корреляциям между родственниками, которые трудно отличить от корреляций, обусловленных генотипическими факторами. Культурная преемственность между родителями и детьми в значительной степени маскирует биологическую наследуемость (Cavalli-Sforza, Feldman, 1974). Это остается одной из самых интересных проблем в биологии.
Возможно, следовало бы также учитывать средовую дисперсию, сопровождающую расовые различия. Например, условия жизни черного и белого населения США сильно различаются. Это может быть обусловлено частично культурными различиями, частично социально-экономическими факторами.
И наконец, существует дисперсия, обусловленная взаимодействием генотипа и среды. У человека такое взаимодействие может иметь место, например, между возрастом наступления половой зрелости и типом школьного обучения, доступным разным возрастным группам.
В общем, мы можем конкретизировать средовую дисперсию, добавляя компоненты, перечисленные выше.
Кавалли-Сфорца и Бодмер (Cavalli-Sforza, Bodmer, 1971) пришли к выводу, что оценки наследуемости не соответствуют относительному значению природы и воспитания; исключения составляют некоторые конкретные обстоятельства. Изменения в условиях среды могут свести на нет даже прямые измерения наследуемости. Например, для особенностей телосложения человека характерна высокая степень наследуемости, однако в популяциях человека эти особенности будут наследоваться по-разному, что обусловлено различными факторами среды, например питанием. Если оценка наследуемости позволяет нам судить об отношении преобладающих генетических различий к преобладающим различиям среды, то эти оценки нельзя относить к другим популяциям, другой среде или другому периоду времени.
biofile.ru
Роль генотипа и условий внешней среды в формировании фенотипа
Роль генотипа и условий внешней среды в формировании фенотипаВсе многообразие всего живого и его постоянное совершенствование были бы невозможны без изменчивости. Это связано с тем, что генотип последовательно реализуется в фенотип в ходе индивидуального развития организма и в определенных условиях среды обитания, факторы которой (колебания освещенности, температуры, влажности, условий питания, взаимоотношений с другими организмами и др.) часто оказывают определяющее значение на проявление и развитие того или иного признака и свойства. Поэтому организмы, имеющие одинаковые генотипы, могут заметно отличаться друг от друга по фенотипу. Приведем несколько примеров.
Если растения примулы, которые в обычных условиях имели красные цветки, перенести в оранжерею и содержать их там при температуре 30—35°С и высокой влажности, то через некоторое время все цветки у них оказываются белыми. Если эти же растения вновь поместить в обычные (комнатные) условия, то распустившиеся цветки будут красными.
Другим классическим примером, иллюстрирующим влияние внешней среды на проявление качественных признаков, служит изменение окраски шерсти у гималайского кролика. Обычно при 20°С у этого кролика шерсть белая, за исключением черных ушей, лап, хвоста и мордочки. При 30°С такие кролики вырастают полностью белыми. Если же у гималайского кролика сбрить шерсть на боку или спине и содержать его при температуре воздуха ниже +2°С, то вместо белой шерсти вырастет черная. Но если сбрить шерсть на ухе, то в обычных условиях там снова вырастет черная шерсть.
Эти наблюдения объясняют, почему гималайские кролики рождаются белыми, без участков черной шерсти: их эмбриональное развитие происходит в условиях высокой температуры.
Известно, что все признаки и свойства организма наследственно детерминированы, однако организмы наследуют не сами признаки и свойства, а лишь возможность их развития. Для проявления и развития признака необходимы соответствующие условия внешней среды.
Но даже в том случае, когда проявление и развитие того или иного признака происходит, степень его выраженности бывает
разной в зависимости от условий внешней среды: при одних она усиливается, при других ослабляется. Пределы этих изменений определены возможностями, заложенными в генотипе.
Пределы модификационной изменчивости признака, ограниченные действием генотипа, называют его норной реакции.
Для разных признаков и свойств организмов границы, определяемые нормой реакции, неодинаковы. Наибольшей пластичностью и изменчивостью характеризуются количественные признаки: семенная продуктивность злаков, величина удоя у крупного рогатого скота, масса животных, число и размеры листьев и колосков у растений и т. д. В то же время качественные признаки (масть животных, окраска семян, цветков и плодов, остистость и опушенность колоса у растений) мало зависят от условий среды.
Так, у крупного рогатого скота удой во многом определяется условиями его содержания. При подборе кормов нужного качества и количества ценная порода скота может дать 5—6 тыс. кг молока в год, а в случае ухудшения условий содержания продуктивность снижается до 2,5 тыс. кг и даже ниже. Что касается масти животных, то при самых разных условиях она почти не изменяется.
Подобную закономерность можно наблюдать и у растений. Например, растения элитного сорта картофеля, выращенные в условиях высокой культуры агротехники, дают урожай клубней 500—600 ц/га и более. Ухудшение условий (тяжелые глинистые почвы, низкая культура земледелия и ухода за растениями) может привести к тому, что урожайность прекрасного сорта картофеля снизится до 100—150 ц/га и ниже. Правда, окраска клубней и их форма при этом изменяются в очень малой степени.
Рассмотренные примеры свидетельствуют о том, что количественные признаки обладают очень широкой нормой реакции, а качественные — более узкой.
Таким образом, все признаки и свойства каждого сорта растений и каждой породы животных могут изменяться только в пределах нормы реакции. Поэтому попытки добиться высоких урожаев, резко повысить продуктивность сельскохозяйственных животных только за счет повышения уровня культуры земледелия, улучшения условий кормления и содержания животных несостоятельны. В первую очередь необходимо получить новые сорта растений и породы животных улучшенного генотипа, отзывчивые на хорошие условия возделывания и выращивания.
По механизму возникновения и характеру изменений признаков различают два типа изменчивости — наследственную и ненаследственную.
Под наследственной изменчивостью понимают способность к изменениям самого генетического материала, а под ненаследственной (модификационной, или фенотипической) — способность организмов реагировать на условия окружающей среды и изменяться в пределах нормы реакции, определяемой генотипом.
Список литературы
Для подготовки данной работы были использованы материалы с сайта http://biology.asvu.ru
coolreferat.com
Генетика поведения животных
Чарлз Дарвин, несомненно, интересовался, существует ли связь между наследственными факторами и поведением, однако пионером в этой области науки был другой внук Эразма Дарвина - Фрэнсис Гальтон. «Происхождение видов» побудило Гальтона посвятить остаток своей жизни изучению наследования психических особенностей. В 1869 г. Гальтон опубликовал книгу «Наследственная гениальность: исследование ее законов и значение». Он считал, что люди выдающихся умственных способностей чаще обнаруживаются среди родственников таких же выдающихся людей, чем среди населения вообще. Не имея какого-либо удовлетворительного метода, позволяющего оценивать умственные способности, Гальтон полагался на показатель репутации, «репутации лидера общественного мнения, изобретателя, человека, перед которым мир осознанно считает себя в большом долгу» (Gallon, 1869). Он проанализировал родословные почти 300 семей, в которых были выдающиеся судьи, государственные деятели, военачальники, писатели, ученые, поэты, музыканты, художники и т. д. Его результаты показали, что видного социального положения с наибольшей вероятностью достигают близкие родственники, а по мере того как родство становится дальше, вероятность стать выдающимся уменьшается.
Гальтон сознавал, что, как правило, выдающиеся люди имеют одинаковые социальные и финансовые преимущества, а также возможности получить образование. Он считал, что репутация выдающихся людей указывает на их природные способности и не обусловлена средовыми факторами. Чтобы обосновать свои аргументы, Гальтон указывал на то, что многие выдающиеся люди произошли из семей ничем особенным не выделяющихся. Он пришел также к выводу, что приемные дети римских католических пап, которые пользуются огромными социальными благами, составляют группу менее известных людей, чем дети выдающихся деятелей. Гальтон затратил значительные усилия, чтобы улучшить способы оценки психических характеристик. Он разработал методики для оценки остроты обоняния, осязания и зрения и для оценки роста и веса, времени реакции и объема памяти. В статистике он ввел понятия корреляции, медианы и процентиля. В 1883 г. он ввел метод исследования близнецов для выяснения относительного влияния генотипа и среды. Он изучил 35 пар идентичных близнецов, которые были абсолютно схожи при рождении и воспитывались в одинаковых условиях. Данные его анализа показали, что их поведенческое сходство сохраняется, даже если во взрослом состоянии они живут раздельно. Галь-тоном были также проанализированы 20 пар неидентичных близнецов, которые были не столь похожи при рождении, но воспитывались в одинаковых условиях. Он писал:
«Никуда не денешься от вывода, что природа чрезвычайно сильно преобладает над воспитанием, когда различия в воспитании не превышают того, что обычно имеет место среди людей одинакового социального положения в одной стране. У меня есть опасения, что мои данные, возможно, доказывают слишком многое и могут восприниматься с недоверием, потому что сильно противоречат житейскому опыту, согласно которому воспитанием можно добиться столь немногого» (Gallon, 1883).
После работ Гальтона стали быстро развиваться исследования генетической основы поведенческих признаков, однако до сих пор в этих исследованиях содержится много противоречий. В начале нашего столетия наибольший интерес вызвал бихевиоризм. В частности, работы Уотсона (Watson, 1930) убедили многих психологов, что развитие поведения не определяется генетическими факторами, и твердые взгляды сторонников влияния средовых факторов на поведение, принятые бихевиористами, оставались господствующими вплоть до шестидесятых годов. Можно сказать, что как наука генетика поведения ведет свое начало с 1960 г., когда Фуллер и Томпсон (Fuller, Thompson, 1960) опубликовали книгу «Генетика поведения». В ней представлена история исследований поведения и умственных способностей человека и рассмотрены данные, свидетельствующие о влиянии генов на поведение. Несмотря на достаточную обоснованность данных, многие социологи и психологи остались противниками идеи влияния генетических факторов на поведение, и споры продолжались. Даже сегодня некоторые теории, основанные на генетической аргументации, остаются противоречивыми. В этой статье мы рассмотрим данные, свидетельствующие о влиянии генетических факторов на поведение.
Единичные гены и поведение
Поведенческие признаки, контролируемые отдельными генами, могут быть предметом строгого генетического анализа поведения. Классический пример этого-работа Маргарет Басток (Margaret Bastock, 1956), изучавшей условия успешного спаривания у плодовой мушки Drosophila melanogaster. Эта исследовательница скрещивала особей, несущих мутацию yellow (желтое тело), с дикими мухами в течение семи поколений. Полученные данные свидетельствовали о том, что дикие мухи генетически сходны с носителями мутации yellow, за исключением участка хромосомы, где локализован ген yellow. Басток обнаружила, что самцы, несущие мутацию yellow, реже спариваются с дикими самками, чем дикие самцы. У желтотелых самцов изменен ритуал ухаживания, и это снижает их успех при размножении. Они в меньшей степени стимулировали самок к спариванию, так как во время ухаживания слишком мало вибрировали крыльями.
Ротенбюлер (Rothenbuhler, 1964) провел тонкий генетический анализ гнездоочистительного поведения медоносной пчелы (Apis mellifera). Личинки этих пчел иногда погибают от болезни, именуемой американской пчелиной гнильцой. Чтобы поддерживать гигиенические условия внутри улья, нормальные рабочие пчелы вскрывают ячейки, в которых находятся больные личинки, и удаляют их. Некоторые расы пчел, называемые «негигиеническими», этого не делают. Скрещенные с нормальными гигиеническими пчелами, они дают негигиенических потомков, что свидетельствует о доминантности этого признака. При скрещивании гибридов с родительской гигиенической расой Ротенбюлер получил следующие результаты. Из 29 образованных колоний в девяти зараженные ячейки оказались вскрытыми, но больные личинки из них удалены не были; в шести - ячейки не вскрывались, но если их вскрывал экспериментатор, то пчелы удаляли больных личинок; в восьми колониях пчелы не вскрывали ячейки и не удаляли личинок.
На основе полученных данных Ротенбюлер предположил, что вскрывание ячеек и удаление личинок контролируется различными генами. Эти результаты можно объяснить, постулируя существование двух пар аллелей, из которых негигиенические аллели доминантны. Таким образом, рабочие пчелы с генотипом Uu или UU не будут вскрывать зараженные ячейки, а с Rr или RR не будут удалять личинок. У гаплоидных самцов пчел, которых называют «трутнями», только один набор хромосом. В 29 семьях Ротенбюлер обнаружил четыре типа трутней (UR, Ur, uR, ur), которых он скрестил с гигиеническими матками (uu, rr), как показано на рис. 4.1. В соответствии с этой простой менделевской схемой возвратное скрещивание дает четыре генотипа в равных соотношениях, и результаты, получетные Ротенбюлером, существенно не отличаются от нее. Среди полностью гигиенических, частично гигиенических или полностью негигиенических рабочих пчел не обнаружено ни физических, ни физиологических различий, хотя имеются данные, что негигиенические пчелы все же могут проявлять гигиеническую активность, но очень редко, и требуют для этого более мощных стимулов, чем нормальные. Это заставляет думать, что аллели U и u действуют как переключатели, которые запускают поведение вскрывания ячеек при условии, что существует некий порог стимуляции.
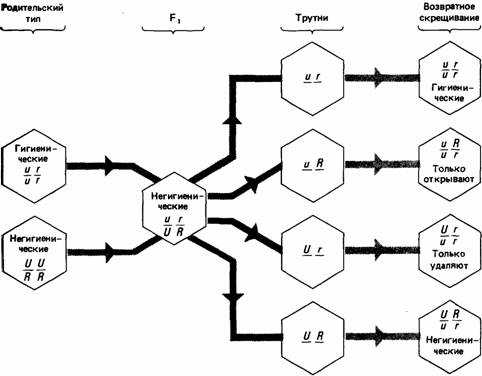
Рис. 4.1. Менделевская схема, предложенная для объяснения устойчивости пчел к американской пчелиной гнильце. (По Rothenbuhler, 1964.)
Гены, которые действуют как переключатели, активируя группу других генов, известны и в ряде других случаев. Например, у некоторых бабочек весьма сложный рисунок крыльев напоминает рисунок крыльев других видов, несъедобных для хищников. Развитие мимикрического рисунка контролируется множественными генами, но оказывается, что определяет появление рисунка единственный ген-переключатель (Sheppard, 1961). Единичные гены известны также и у человека. Примером может служить недостаточность лак-тазы, обнаруженная у некоторых расовых групп. Синтез лактазы контролируется геном с тремя аллелями - L, l 1 и l 2. Как l 1, так и l 2 рецессивны по отношению к L, а l 2 рецессивен по отношению к l 1. У особей с генотипами LL, Ll 1 или Ll 2 лактаза синтезируется как у взрослых, так и у детей. Особи с генотипом l 1l 1 или l 1l 2 не синтезируют лактазу во взрослом состоянии, а особи с l 2l 2 не могут ее синтезировать даже в детстве. Взрослые люди с генотипами l 1l 1 или l 1l 2 могут употреблять в пищу только кисломолочные продукты.
Хромосомные мутации
Расположение и число хромосом часто можно наблюдать непосредственно под микроскопом. Известны различные типы хромосомных мутации и влияние некоторых из них на фенотип. Поэтому изучение связи между поведением и строением хромосом представляет собой вполне подходящий метод для исследования влияний генов на поведение.
Излюбленным объектом такого типа исследований является плодовая мушка Drosophila. В клетках слюнных желез личинок дрозофилы содержатся гигантские хромосомы; препараты слюнных желез сравнительно легко приготовить и анализировать. Впервые хромосомный анализ при изучении поведения дрозофилы применили Хирш (Hirsch) и его коллеги. Они изучали стремление Drosophila melanogas-ter двигаться в направлении силы земного притяжения или против него (положительный или отрицательный геотаксис). Поведенческую реакцию они проверяли в вертикальном пластиковом лабиринте (рис. 4.2), который должны были преодолеть мухи, привлекаемые запахом пищи.
Обратному движению препятствовали конусообразные воронки в соединениях лабиринта. Большое число мух запускали в пробирку с левой стороны лабиринта и собирали в серию пробирок справа. Таким способом можно проверить тысячи мух, не трогая их при этом руками. Среди мух, собранных в различные пробирки с правой стороны, можно было выделить мух со строго положительным геотаксисом, со строго отрицательным геотаксисом и промежуточных. Скрещивая мух из крайней верхней и крайней нижней пробирок, можно получить расы с положительным или отрицательным геотаксисом (рис. 4.3).
В одном из экспериментов сравнивали три популяции дрозофил (Hirsch, Erlen-meyer-Kimling, 1962). В одной вели отбор на положительный геотаксис, в другой - на отрицательный геотаксис, а в третьей, контрольной, популяции проводилось неселективное скрещивание. Мух скрещивали с особой линией, в которой были различные хромосомные инверсии и гены-маркеры. У Drosophila melanogaster имеются четыре пары хромосом, три пары больших и одна - маленьких. Гены-маркеры использовались для идентификации трех больших хромосом. Это были доминантные гены, контролирующие признаки, отчетливо проявляющиеся в фенотипе. Специальная схема скрещивания позволила получить самок, которые были либо гомозиготны, либо гетерозиготны по хромосомам, подлежащим исследованию.
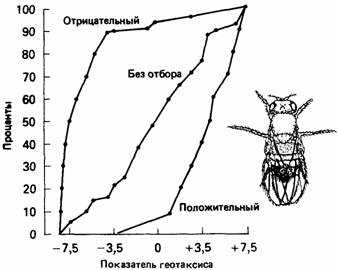
Рис. 4.3. Показатели, полученные при изучении геотаксиса, у трех линий Drosophila в лабиринте.
Эти три хромосомы можно было идентифицировать с помощью генов-маркеров следующим образом: хромосома Х-ген узких глаз (Bar), хромосома II-ген волнистых крыльев (Cy) и хромосома III - ген, обусловливающий короткие щетинки (Sb). Тестируемую самку, несущую эти маркеры, скрещивали с самцами одной из исследуемых линий. Для использования в последующих экспериментах из потомства отбирали только тех, которые несли все три гена-маркера. При возвратных скрещиваниях, используя самцов из исходной популяции, получили восемь возможных генотипов. Следовательно, каждая из трех больших хромосом гетерозиготна или гомозиготна по S-хромосоме, полученной от тестируемой линии (s), к которой принадлежал отец. На генотипах этих восьми классов изучали индивидуальные эффекты и взаимодействие хромосом.
Хирш и Линда Эрленмейер-Кимлинг (Hirsch, Erlenmeyer-Kimling, 1962) обнаружили, что в популяции, где отбор не проводился, хромосомы X и II оказывают большое влияние на развитие положительного геотаксиса, а хромосома III-на развитие отрицательного геотаксиса. В линии, где проводился отбор на положительный геотаксис, были обнаружены небольшие изменения в хромосомах X и II, но хромосома III в этом случае оказывала влияние на развитие положительного геотаксиса. В линии, которая была отобрана на отрицательный геотаксис, отрицательное влияние хромосомы III увеличивалось, тогда как положительное действие хромосом X и II было меньше. Эффект от совместного действия всех трех хромосом был больше в случае отрицательного геотаксиса. Это неудивительно, так как общая реакция на отбор сильнее в случае отрицательного геотаксиса. Эти результаты показывают, что поведение, связанное с геотаксисом, контролируется рядом генов, которые находятся во всех трех больших хромосомах.
Хромосомный анализ широко применяется при изучении различных типов поведения дрозофилы, в том числе скорости спаривания и других аспектов ухаживания (Ehrman, Parsons, 1976). Хромосомные инверсии обычны, и обнаружено, что D. pseudoobscura с такими инверсиями в гетерозиготном состоянии характеризуются большей приспособленностью, чем с ин-.версиями в гомозиготном состоянии, благодаря влиянию на поведение ухаживания.
Рис. 4.4. Структура фразы призывного сигнала сверчка Teleogryllus. Каждая фраза состоит из пульсов лвух типов: пульсы типа А содержатся в составе серии, а пульсы типа В - в составе трели. Песня Т. oceanicus показана вверху, а песня Т. commodus - внизу. (По Bentley, Hoy, 1972.)
Хромосомные аномалии у человека являются предметом интенсивных исследований, и считают, что эти аномалии являются причиной таких заболеваний, как эпилепсия, маниакально-депрессивный психоз, умственная отсталость и шизофрения (McClearn, DeFries, 1973; Ehrman, Parsons, 1976).
Полигенное наследование поведения
Многие поведенческие признаки контролируются большим числом генов и большим числом факторов окружающей среды. Для генетического анализа таких сложных ситуаций можно использовать разнообразные методы. Их эффективность в большой мере зависит от степени наследуемости рассматриваемого поведенческого признака. Однако некоторые аспекты поведения характеризуются высокой степенью наследуемости, с них мы и начнем обсуждение полигенного наследования.
Самцы сверчков привлекают самок с большого расстояния призывными сигналами. Звук извлекается при ритмичном раскрывании и складывании специализированных надкрылий, на которых находится специальный фрикционный механизм. При каждом складывании надкрылий возникает звук, тогда как раскрывание беззвучно. Эти призывные сигналы удивительно стереотипны у членов локальной популяции, но они резко различаются у разных видов. Различия имеют место в основном во временной организации звуковых пульсов (рис. 4.4). Межвидовые гибриды в природе не встречаются (Hill et al., 1972), однако их можно получить в лаборатории. Например, Леруа (Leroy, 1964) получил гибрид между австралийскими полевыми сверчками Teleogryllus commodus и Т. oceanicus. Бентли и Хой (Bentley, Hoy, 1972) обнаружили, что призывные сигналы гибридов первого поколения (Fj) заметно отличаются от призывных сигналов обоих этих видов. В частности, интервалы между сериями и между трелями гибридов имели значения, промежуточные между значениями соответствующих интервалов у родителей. На рис. 4.5 сравниваются осциллограммы призывных сигналов Т. commodus и их гибридов. По данным Бентли и Хоя, реципрокные гибриды отличаются друг от друга. Призывный сигнал гибрида, полученного от самки Т. oceanicus и самца Т. commodus, был похож на сигнал Т. oceanicus наличием отчетливо выраженного интервала между трелями. У гибрида, полученного от самки Т. commodus и самца Т. oceanicus, в звуковом сигнале отсутствует хорошо выраженный интервал между трелями, и он похож на сигнал Т. commodus. Эти различия лучше всего видны на гистограмме распределения межимпульсных интервалов (рис. 4.6.).
Эти результаты позволяют предположить, что наследование характера при зывного сигнала полигенно. Мы не располагаем данными о том, что какой-либо конкретный признак призывного сигнала является доминантным и призывные сигналы гибридов промежуточны по своим параметрам между призывными сигналами родителей.
Рис. 4.5. Осциллограммы призывных сигналов Т. oceanicus (А.). Т. commodus (Г) и их гибридов: самка Т. oceanicus х самец Т. commodus (Б) и самка Т. commodus х самец Т. oceanicus (В) (стрелками отмечено начало фраз). (По Bentley, Hoy, 1972.)
Эта гипотеза подкрепляется тем фактом, что у гибридов от возвратного скрещивания также наблюдается промежуточное наследование (Bentley, 1971). Кроме того, существуют некоторые указания на то, что наследование этих признаков сцеплено с полом. Призывные сигналы гибридов от реципрокного скрещивания отличаются таким образом, что можно думать, что интервалы между трелями, свойственные Т. oceanicus, есть у одних гибридов, но отсутствуют у других. Обозначение пола у сверчков, ХО, означает, что Y-хромосома отсутствует. Самец получает Х-хромосому от матери, но не получает ничего, что соответствовало бы ей от отца. Таким образом, оказывается, что интервал между трелями у гибрида от самки Т. oceanicus и самца Т. commodus определяется Х-хромосомой матери Т. oceanicus. Этого признака нет ни у гибридов от самки Т. commodus и самца Т. oceanicus, ни у их матерей.
На призывные сигналы самцов различных видов реагируют преимущественно самки соответствующего вида. Интересно, однако, узнать, как гибридные самки реагируют на призывные сигналы своих родителей и своих братьев. Самки сверчков немые и реагируют на призывные сигналы самца тем, что ползут по направлению к источнику звука. Хилл и др. (Hill et al., 1972) показали, что самки Т. oceanicus и Т. commodus, имеющие возможность свободно перемещаться, могут отличать призывный сигнал своего вида от призывного сигнала других видов.
Чтобы выразить количественно предпочтение самки, Хой и Пол (Ноу, Poul, 1973) использовали Y-образный лабиринт. Они воспроизводили записанные призывные сигналы через громкоговоритель, помещенный справа и слева от лабиринта. Таким способом они имели возможность измерять относительную привлекательность любого призывного сигнала. Хой и Пол обнаружили, что самки Т. oceanicus и Т. commodus отличают видовой призывный сигнал; это же было обнаружено Хиллом и др. у свободно перемещающихся самок. Кроме того, эти авторы показали, что гибридные самки, являющиеся дочерьми самок Т. oceanicus и самцов Т. commodus, призывный сигнал своих братьев предпочитают типичному призывному сигналу любого из родителей. Как видно из рис. 4.6, призывный сигнал гибрида от скрещивания самца Т. oceanicus и самки Т. commodus легко отличить от призывного сигнала любого из родителей. (Обратите внимание, что он не так четко разделен на интервалы, как в случае с гибридом от другой комбинации.) Тот факт, что призывный сигнал гибрида предпочитается самкой, означает, что призывный сигнал самца и восприятие и распознавание его самкой каким-то образом связаны генетически.
Рис. 4.6. Гистограммы частот интервалов между пульсами в призывных сигналах сверчков. Каждая гистограмма представляет собой анализ одного индивидуального сигнала. На гистограммах можно различить внутрисерийные интервалы, интервалы в пределах трели и интервалы между трелями. (По Bentley, Hoy, 1972.)
Хорошо известно, что различные породы собак характеризуются разными поведенческими признаками. В 1965 г. Скотт и Фуллер (Scott, Fuller, 1965) опубликовали некоторые результаты своих обширных исследований по генетической основе таких различий. В одном исследовании они сравнивали поведение коккер-спаниелей, басенджи и их гибридов. Спаниели с давних пор воспитывались как спортивные собаки, послушные и преданные своему хозяину. Родиной спаниелей считается Испания, где их использовали на охоте с ловчими птицами и при ловле птиц сетями. Собаку приучали прижиматься к земле, когда она обнаружит куропатку или другую дичь. Затем брошенной сетью накрывали и птицу, и собаку. После изобретения огнестрельного оружия сеть больше не использовали и спаниелей научили останавливаться и указывать, где они обнаружили птиц. Были выведены спрингер-спаниели, которые прыгали и вспугивали птиц. Первоначальная привычка затаиваться осталась только у коккер-спаниелей.
Басенджи широко распространены в Африке; пигмеи и некоторые другие африканские племена используют их на охоте. Это многоцелевые охотничьи собаки, довольно осторожные. Басенджи молчаливы и почти никогда не лают, хотя порой они долго и протяжно воют.
Скотт и Фуллер (Scott, Fuller, 1965) осуществили обширную программу скрещивания этих двух пород (рис. 4.10), включая реципрокные скрещивания и различные типы возвратного скрещивания. В табл. 4.1 суммированы признаки этих двух пород и указаны наиболее вероятные способы наследования, основанные на результатах экспериментов. Щенки басенджи обычно дикие в сравнении с ручными и ласковыми щенками спаниелей. Они избегают рук и сопротивляются, если их пытаются удержать. Проверка на приручение показала, что поведение щенков F1 сходно с поведением родителей и определяется, таким образом, доминантными генами. Результаты экспериментов по возвратному скрещиванию свидетельствуют о том, что единичный доминантный ген контролирует дикость, обычно свойственную басенджи. Способность к послушанию у спаниелей контролируется единичным рецессивным геном. Возьмем другой пример. У самок басенджи течка бывает обычно раз в год, во время осеннего равноденствия. У спаниелей, как и у большинства европейских пород, это происходит каждые шесть месяцев в любое время года. Эстральный цикл у басенджи, очевидно, контролируется единичным рецессивным геном. Учитывая все это, Скотт и Фуллер пришли к выводу, что исследуемые ими поведенческие признаки контролируются одним или двумя генами. Эта ситуация оказывается промежуточной между простым менделевским наследованием и полигенным наследованием.
Таблица 4.1.
|
Признаки |
Бассенджи |
Коккер-спаниели |
Наиболее вероятный способ наследования |
|
Избегание и голосовые реакции при взятии на руки |
Высокое |
Низкое |
Один доминантный ген, определяющий дикость |
|
Сопротивление ограничению движений |
Высокое |
Низкое |
Один ген без доминирования |
|
Агрессивность во время игры в возрасте 13-15 нед |
Высокая |
Низкая |
Два гена без доминирования |
|
Порог стимуляции |
Высокий |
Низкий |
Два доминантных гена, определяющие низкий порог |
|
Тенденция лаять малое число раз |
Высокая |
Низкая |
Один ген без доминирования |
|
Половое поведение (время течки) |
1 раз в год |
1 раз в полгода |
Тип басенджи определяется одним рецессивным геном |
|
Тенденция сохранять спокойствие при взвешивании |
Низкая |
Высокая |
Два рецессивных гена, определяющих сильную тенденцию. |
Может показаться удивительным, что такие сложные черты поведения контролируются таким малым числом генов. Однако эти две породы собак были изолированы в течение долгого времени и подвергались интенсивному искусственному отбору. Это, по-видимому, сформировало генотип, который гомозиготен по отдельным признакам, так что имеется лишь небольшое расщепление внутри пород. Генетика поведения собак особенно интересна, так как способность научиться конкретному типу поведения у них часто является наследственной.
Рис. 4.10. Родительские пары коккер-спаниелей (вверху) и басенджи (внизу), использованные в генетических исследованиях Скотта и Фуллера. (Skott, Fuller, 1965; фотография J. P. Scott.)
Наследование поведенческих признаков
«Вопрос о том, что важнее в формировании физических черт человека и его личности - природа или воспитание, генотип или окружающая среда, - попросту ошибочен и может ввести в заблуждение. Генотип и окружающая среда важны в равной степени, так как и то и другое необходимо... Вопрос о роли генотипа и окружающей среды в развитии человека следует поставить так: до какой степени различия, наблюдаемые между людьми, определяются различиями их генотипов и различиями условий, в которых люди рождаются, растут и воспитываются? (Dobzhansky, 1964).»
Большая часть работ Фрэнсиса Гальтона основана на представлении о том, что фенотипическое сходство между родственниками частично определяется сходством их генотипов. Однако Гальтон не был знаком с работами Менделя и считал, что наследственность носит слитный характер. Он думал, что материал наследственности непрерывен, поэтому признаки потомков промежуточны между признаками их родителей. На основе таких допущений Гальтон впервые разработал методику расчета корреляций между родственниками как средство оценки наследования тех или иных признаков. В 1918 г. Фишер опубликовал статью, в которой было показано, что возможно предсказать корреляции, ожидаемые между родственниками на основе теории Менделя. Работы Менделя стали известны общественности в 1900 г., однако они относились только к дискретным признакам. Фишер попытался применить менделевскую теорию для анализа непрерывных признаков, таких, как рост и масса. Интересно, что его выводы были сходны с выводами Гальтона. причем большая часть прогнозов в этих двух теориях была почти идентична.
Наследуемость впервые была определена как доля изменчивости, связанная с различиями в наследственности (Lush, 1940). Иными словами, когда мы анализируем конкретный признак в популяции животных, такой, например, как масса тела, мы определяем, какой вклад вносят в фенотипическое проявление этого признака генотипическая и средовая компоненты. Рассматривая родственные популяции в различных условиях, иногда можно оценить степень влияния на изменчивость особей факторов окружающей среды и генетических факторов.
Существуют разнообразные статистические методы оценки наследуемости поведения. Некоторые основаны на исследовании фенотипической изменчивости у генотипически однородных особей. Эта общая фенотипическая изменчивость генотипически идентичных особей сравнивается с общей фенотипической изменчивостью в природной генотипически разнообразной популяции. Такое сравнение дает соотношение, в котором противопоставляется генотипическая компонента фенотипического разнообразия средовой компоненте. При оценке с помощью этой статистической методики мы сталкиваемся с определенными трудностями, но она ясно показывает, насколько ошибочно относить один вид поведения к «наследственному», а другой - к «приобретенному». В случае если возникнет новая генная вариация или популяция попадает в другие условия среды, это соотношение может сильно измениться. Типы поведения, которые «генетически детерминированы» в одной среде (у монозиготных близнецов, воспитанных вместе, фенотипы оказываются сходными), могут быть подвержены и сильному влиянию средовых факторов, например климатических (у монозиготных близнецов, воспитанных порознь, фенотипы оказываются разными). Напротив, тип поведения, который «детерминирован средовыми факторами» (у всех генотипически различающихся особей одинаковый фенотип, если их воспитывают в одинаковых условиях), может, очевидно, в значительной степени подвергаться воздействию генетической компоненты в среде, в которой проявляется генотипическое разнообразие. Все эти взаимодействия позволяют лишь в строгих пределах определить степень наследуемости того или иного типа поведения, а также провести границу между врожденным и приобретенным поведением. Мы можем сказать, что данный тип поведения может наследоваться в 80% случаев при данном генотипе, в данной среде и в конкретной популяции, но мы не можем переносить эти выводы на другую популяцию или среду.
При оценке наследуемости ошибки могут проистекать из разных источников, наиболее важный из них - это взаимодействие генотипа и среды и корреляции генотипа и среды. Взаимодействие между генотипом и средой вводит изменчивость, которая не принимается во внимание при обычных расчетах наследуемости. Например, Трайон (Тгуоп, 1942) проводил селекцию крыс на неспособность ориентироваться в лабиринте («глупые» крысы) и на блестящее ориентирование в нем («умные» крысы). Этих крыс выводили и тестировали в обычных лабораторных условиях. А Купер и Зубек (Cooper, Zubek, 1958) выращивали одних крыс в обычных условиях, других - в обедненных, без укрытий и препятствий, а третьих - в условиях, обогащенных сложными лабиринтообразными сооружениями. При испытаниях в стандартном лабиринте эти две линии крыс, выращенные в обычных условиях, сильно различались по своим возможностям ориентироваться в лабиринте, как это обнаружили раньше и другие исследователи. Однако обе линии крыс, выращенных в обедненных условиях, одинаково плохо ориентировались в лабиринте, тогда как обе линии, выращенные в обогащенной среде, ориентировались одинаково хорошо. Хотя эти две линии отчетливо различаются генотипически, в фенотипе эта разница проявляется только у крыс, выращенных в определенных условиях среды.
Корреляция генотипа и среды - другой потенциальный источник ошибок при оценке наследуемости поведенческих признаков - может возрастать, когда особи избирают какую-то особую среду или у них развиваются определенные привычки для компенсации генотипических дефектов. Однако, несмотря на эти трудности, оценка наследуемости оказалась особенно полезной для разделения эффектов природы и воспитания.
Оценки наследуемости, основанные на сходстве между родственниками, часто использовались в генетическом анализе поведения человека. Однако здесь есть ряд осложняющих факторов. Во-первых, генетическое доминирование снижает корреляцию между родственниками. При генетическом доминировании вклады родителей в проявление признака неодинаковы, т. е. вводится еще один источник изменчивости. Это называется дисперсией доминирования. Однако дисперсия доминирования оказывает различное действие на корреляции родителей и потомков и корреляции между сибсами. Фактически разница между этими двумя коэффициентами корреляции должна точно составлять одну четверть дисперсии, обусловленной доминированием (Bodmer, Cavalli-Sforza, 1976).
Во-вторых, подбор пар у человека, вероятно, не является случайным в отношении некоторых признаков. Можно допустить, что в отношении определенной части признаков, которые нельзя наблюдать непосредственно, например уровней ферментов, подбор пар происходит случайным образом, однако этого нельзя сказать в отношении таких признаков, как рост и коэффициент умственного развития. Корреляции между мужем и женой по этим признакам обычно положительны, что указывает на неслучайный или выборочный подбор, поскольку люди обычно стремятся выбирать партнеров одинакового роста и умственного развития. Эти корреляции по росту составляют примерно +0,3, а по умственному развитию - примерно + 0,4. На эти корреляции могут влиять и социальные факторы, так как между социально-экономическим положением и данными признаками также существует положительная корреляция. Поэтому тенденция выбирать мужа или жену из той же социальной группы обычно приводит к положительной корреляции между мужем и женой. Ассортативные браки у человека могут влиять на увеличение частоты гомозигот.
В-третьих, при анализе наследуемости поведения человека необходимо правильно оценивать средовую дисперсию, поскольку в этом случае условия среды нельзя контролировать, как при исследованиях, проводимых на животных. Был предложен способ (Cavalli-Sforza, Bodmer, 1971) разделения средовой дисперсии на отдельные компоненты. Прежде всего следует различать внутрисемейную дисперсию. Она может быть обусловлена возрастными различиями, порядком рождения, различиями по полу и т.д. В семьях разных размеров эта дисперсия может также зависеть от уровня питания и наличия жилищной площади.
Далее существует дисперсия, обусловленная различиями между семьями одного социально-экономического слоя. Некоторую информацию о значении этого фактора можно почерпнуть из рассмотрения корреляции между приемными детьми и их приемными родителями, однако результаты могут быть тенденциозны из-за того, что родителей выбирают посреднические агентства.
Затем следует учитывать дисперсию между социально-экономическими слоями. Культурная преемственность может привести к корреляциям между родственниками, которые трудно отличить от корреляций, обусловленных генотипическими факторами. Культурная преемственность между родителями и детьми в значительной степени маскирует биологическую наследуемость (Cavalli-Sforza, Feldman, 1974). Это остается одной из самых интересных проблем в биологии.
Возможно, следовало бы также учитывать средовую дисперсию, сопровождающую расовые различия. Например, условия жизни черного и белого населения США сильно различаются. Это может быть обусловлено частично культурными различиями, частично социально-экономическими факторами.
И наконец, существует дисперсия, обусловленная взаимодействием генотипа и среды. У человека такое взаимодействие может иметь место, например, между возрастом наступления половой зрелости и типом школьного обучения, доступным разным возрастным группам.
В общем, мы можем конкретизировать средовую дисперсию, добавляя компоненты, перечисленные выше.
Кавалли-Сфорца и Бодмер (Cavalli-Sforza, Bodmer, 1971) пришли к выводу, что оценки наследуемости не соответствуют относительному значению природы и воспитания; исключения составляют некоторые конкретные обстоятельства. Изменения в условиях среды могут свести на нет даже прямые измерения наследуемости. Например, для особенностей телосложения человека характерна высокая степень наследуемости, однако в популяциях человека эти особенности будут наследоваться по-разному, что обусловлено различными факторами среды, например питанием. Если оценка наследуемости позволяет нам судить об отношении преобладающих генетических различий к преобладающим различиям среды, то эти оценки нельзя относить к другим популяциям, другой среде или другому периоду времени.
biofile.ru