Часть I. Общая характеристика психической деятельности животных. Наблюдение поведенческих реакций животных на факторы внешней среды
14. Внутренние факторы инстинктивного поведения животных.
Внутренние факторы инстинктивного поведения. На протяжении длительного времени считалось, что научение определяется внешними факторами, а инстинктивное поведение – исключительно внутренними факторами, причем природа этих факторов была неизвестна. Поиск, уточнение внутренних факторов инстинктивного поведения позволили бы ответить на вопрос, что же определяет мотивацию поведения.
Внутренние факторы, бесспорно, оказывают влияние на инстинктивное поведение животных. В середине XX в. американский биолог П. Уит проводил опыты с пауками, в ходе которых изучал плетение паутины при попадании в организм животного разных химических веществ. Нужное вещество наносилось в виде капли непосредственно на паутину или вводилось при помощи шприца в попавшуюся жертву. Каждое вещество стимулировало паука плести паутину определенного вида, сама же реакция плетения паутины является у паука наследственной. Так, кофеин заставлял пауков плести бесформенную паутину из беспорядочно спутанных нитей, паук при этом испытывал подобие невроза. При поступлении в организм первитина паук становился очень беспокойным и не сплетал паутину целиком. Хлоргидрат вызывал у паука оцепенение, и он не доплетал паутину. А лизергиновая кислота способствовала повышению концентрации внимания на плетении, и паук сплетал паутину очень тщательно и ровно, при этом ее качество превосходило естественное.
Внутренняя среда организма постоянна, разнообразные процессы регуляции направлены на поддержание физико-химического состава среды. Она постоянно обновляется, однако все ее параметры поддерживаются на определенном уровне за счет саморегуляции, что обеспечивает протекание всех биохимических реакций. Особенность внутренних процессов животного организма состоит в том, что они часто протекают в форме ритмов. В 1930-е гг. советский зоопсихолог В.М. Боровский выдвинул предположение о том, что именно отклонения этих внутренних ритмов организма от нормы являются первичной мотивацией поведенческих реакций. В определенных условиях внутренняя согласованность физиологических ритмов нарушается, и прежнее равновесие в новых условиях не обеспечивает нормальное функционирование организма. Возникает внутреннее побуждение, направленное на восстановление внутреннего равновесия, т. е. появляется потребность. Инстинктивное поведение в этом случае будет направлено на удовлетворение этой потребности.
Важнейшими источниками внутренних стимулов инстинктивного поведения являются гормоны и рецепторы. Известно, что половые гормоны и гормоны гипофиза стимулируют целый ряд форм поведения, связанных с размножением, – драки между самцами за самку и за территорию, охрана гнезда, брачные игры.
Для внутренней мотивации большое значение имеют прежде всего ритмические процессы, которые происходят в центральной нервной системе. Ритмическая активность ее стволовой части у позвоночных животных и брюшных нервных структур у беспозвоночных обеспечивает ориентацию поведения во времени. Известно существование у животных так называемых «биологических часов» – автономных колебательных процессов, которые осуществляют регуляцию всех ритмов жизнедеятельности организма. «Биологические часы» определяют колебания внешней активности поведения животных, все действия, которые повторяются с определенной цикличностью. Они как бы закладывают основу инстинктивного поведения животного, а факторы среды вносят в эти ритмы свои коррективы. Изменения могут быть связаны с действием разнообразных внешних стимулов (слуховых, зрительных и др.), а также могут зависеть от общего физиологического состояния животного в данный момент. Чаще всего в поведении животных отмечаются циркадные, или суточные, ритмы, период которых равен суткам.
Интересно отметить, что активность животного подвержена таким ритмическим суточным колебаниям даже в условиях полной изоляции от всех факторов естественной среды. Например, животное можно поместить в условия полного круглосуточного освещения и тем не менее наблюдать у него чередование периодов сна и бодрствования, близкое к естественным. Кроме того, в течение дня в поведении животных могут быть отмечены и краткосрочные ритмы. Примером могут послужить наблюдения немецкого этолога В. Шлейдта за индюками. Он отметил, что клохтанье индюка в течение дня повторяется с определенным ритмом, который сохраняется даже при полной изоляции птицы и лишении ее слуха.
Помимо ориентации поведения животного во времени, «биологические часы» ориентируют его в пространстве. Например, перелетные птицы при ориентации по положению солнца должны в каждый момент времени соотносить его положение со временем дня. Это происходит, когда они соотносят информацию о положении солнца с внутренними циркадными ритмами.
Внутренние факторы создают в организме состояние, предшествующее проявлению той или иной инстинктивной реакции. Однако начало этой реакции может зависеть от внешних условий среды. Например, определенный уровень половых гормонов и гормонов гипофиза стимулирует различные поведенческие реакции животного, связанные с размножением, но продуцирование этих гормонов приурочено к определенному времени года. Если животное, которое обитает в умеренной зоне Северного полушария, с наступлением весны содержать в условиях короткого светового дня, активность желез проявляться не будет. Напротив, если зимой создать животному условия постепенно увеличивающегося дня, гормоны начнут выделяться, и половое поведение проявится в зимнее время года.
Внутренние факторы обеспечивают готовность организма выполнить то или иное инстинктивное движение, внешние стимулы могут быть не обязательны для проявления инстинктивной реакции.
Немецкий нейрофизиолог Э. Хольст обнаружил в стволе головного мозга курицы несколько зон. При воздействии на эти зоны слабым электрическим током возникают инстинктивные движения, соответствующие той или иной зоне. Было отмечено, что если длительное время воздействовать на одну и ту же зону, усиливая силу раздражения, можно наблюдать целый ряд инстинктивных действий, которые будут совершаться в том же порядке, что и в естественных условиях. Например, курица выполняла движения, которые совершаются при приближении наземного хищника: проявляла легкое беспокойство, потом поднималась, хлопала крыльями, вскрикивала, а затем взлетала. При этом самого раздражителя (хищника) в пределах ее видимости не было. Таким образом, под влиянием исключительно внутренних факторов могут проявиться не только отдельные инстинктивные движения, но и целые инстинктивные действия. Однако при этом не следует забывать, что в естественных условиях инстинктивные действия «запускаются» все же внешними факторами. Приближение наземного хищника, которого увидела бы курица, привело бы к возбуждению соответствующей зоны головного мозга птицы, которая в условиях опыта была простимулирована искусственно.
studfiles.net
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ПОВЕДЕНИЕ — МегаЛекции
Поведение животных формируется и проявляется в зависимости от состояния внутренней среды организма и внешних воздействий и постоянно изменяется. В одних случаях это помогает приспособиться к меняющимся условиям жизни, а в других — может носить негативный характер и привести к снижению адаптации животных и заболеваниям.
Более изменчивы приобретенные реакции и сформировавшиеся условные рефлексы. Инстинкты у животных очень прочные и наследственно закреплены, но и они иногда тоже изменяются. Полезные для данной популяции изменения врожденного поведения могут в дальнейшем передаваться потомству. Поэтому основные формы поведения животных эволюционируют вместе с видом.
Что же может оказать влияние на поведение животных?
Наследственность.Наблюдения за животными-близнецами, помещенными в различные условия, выявили сходство их основных поведенческих реакций: практически одинаковая скорость образования, дифференцировки и переделки условных рефлексов, а также таких качеств, как смелость, трусость, упрямство, отношение к человеку.
Хорошо известны породные отличия в поведении животных. Например, доберман-пинчеры характеризуются повышенной воз-
будимостью, а кавказские овчарки —флегма: ичп<1( м.и I v \» рчи вых лошадей преобладают сильные, подвижные типы ИНД, и у ш желовозов — инертные.
Наследственные факторы определяют те формы поведении, кп торые базируются на безусловных рефлексах (половая активное 11., агрессивность, драчливость или же уклонение от внутри груш ю вых конфликтов). Поэтому селекция животных по показателям типов ВНД, рангов животных, их продуктивности, стрессоустойчивое™ может быть эффективной для формирования стада молочных коров, пригодных к условиям машинного доения.
В процессе одомашнивания велся отбор животных в желательном для человека направлении, что сопровождалось и изменениями условий жизни. Поэтому поведение домашних животных существенно отличается от их диких предков. Некоторые инстинкты у них исчезли — миграция у уток и гусей, насиживание у кур-несушек, агрессивность по отношению к человеку. Тем не менее многие виды биологического поведения у домашних животных сохранились, они проявляются в стаде, на пастбище, в просторных загонах—установление иерархических отношений, характерные движения во время кормления, в период половой охоты и др.
Типы высшей нервной деятельности (ВНД).Животные разного типа ВНД неадекватно проявляют себя в одинаковых ситуациях. Так, животные-лидеры обычно имеют сильный уравновешенный тип, а низших социальных рангов — слабый тип нервной системы.
От типа ВНД зависит быстрота выработки и прочность закрепления условных рефлексов, динамического стереотипа, устойчивость к неблагоприятным условиям содержания.
Условия окружающей среды.Поведение направлено на удовлетворение биологических потребностей животного (питание, воспроизводство, движение, игра и т. п.) и обусловлено изменениями окружающей среды. Например, меняются формы пищевого, полового, социального поведения у коров во время смены стойлового содержания на пастбищное.
Периодические сезонные изменения среды (освещенность, солнечная радиация, климат, состав кормов и пр.) вызывают повторяемость определенных физиологических процессов, их качественные и количественные особенности. Эти изменения называются сезонными ритмами. Примером может служить сезонный ритм воспроизводительных функций. У животных очень четко проявляются и суточные (циркадные) ритмы, связанные с вращением Земли и продолжительностью светового дня, такие, как чередование сна и бодрствования, подвижность, колебания вегетативных процессов — артериального давления, температуры тела, концентрации гормонов в крови и многое другое. Формирование общего суточного ритма поведения животных зависит от установленного распорядка дня, в частности от кратности кормления и доения.
Знание периодичности сезонного и суточного поведения животных используется в практике животноводства для повышения продуктивности. Так, искусственное увеличение продолжительности светового дня в птичнике продлевает цикл яйцекладки у кур.
Одной из причин, нарушающих поведение животных, является шум от работающих в помещении механизмов. При уровне шума 40...60 дБ животные, особенно лошади и коровы, проявляют беспокойство, у них понижаются аппетит и поедаемость кормов, сокращается время отдыха. Но к такому уровню шума они привыкают, и биологическое поведение восстанавливается. Однако при возрастании шума до 60...70 дБ продуктивность снова падает.
Изменяется поведение животных при перегруппировках. При вводе в группу новичка вначале с ним «выясняют отношения» животные высших рангов, а остальные выжидают. Чужака окружают, вступают с ним в единоборство и стараются изгнать из группы, особенно если это животное высокого ранга. При введении в группу животного низкого ранга реакция группы более слабая и непродолжительная. Вводимые животные также по-разному реагируют на новую обстановку: низкого ранга сразу признают свое поражение, а высшего принимают вызов, при этом частота дыхания у них доходит до 100 при пульсе более 100 ударов в 1 мин. Отделение от группы часто вызывает стрессовое состояние. Животное, оставленное в загоне, когда остальных уводят, проявляет сильное беспокойство, стремится соединиться с группой. Известно, что разлученные коровы или лошади, ранее симпатизировавшие друг другу, нередко похожи на больных животных: у них пропадает аппетит, отмечаются вялость, депрессия, угнетенное состояние.
При резких изменениях питания, климата, продолжительности светового дня поведение животных меняется с целью наилучшего приспособления организма к новым условиям. Так, завезенные из Канады в Сибирь бобры успели за лето построить и оборудовать жилище и запасти на предстоящую зиму корм — ветки и стволы деревьев. Однако сибирская зима более длительная и суровая, чем на их родине, и в первой зимовке им не хватило запасенного корма, но на следующий год семья бобров заготовила значительно больше корма и прекрасно перезимовала.
Сельскохозяйственные животные в условиях круглогодового стойлового содержания находятся среди относительно постоянных факторов окружающей среды (температура и влажность помещения, освещенность, гиподинамия, состав корма, одни и те же окружающие животные) и приспосабливаются к ним соответствующим образом, т. е. у них вырабатывается определенный стереотип поведения. Однако за адаптацию к слишком постоянным условиям жизни организм расплачивается потерей пластичности и способности успешно противодействовать неблагоприятным факторам. Поэтому периодическое изменение микроклимата, светового дня, а также прогулки, пастьба, игры оказывают тренирую-
щее воздействие на многие системы организма, что способствует лучшему физиологическому развитию и разнообразию жизненных проявлений, повышению резистентности к болезнетворным факторам. В то же время частые и интенсивные вмешательства в естественный жизненный ритм животного могут вызвать неблагоприятные физиологические реакции, нарушение адаптации и стрессовое состояние.
Физиологическое состояние животных.Голод и сытость, беременность и лактация, послеродовой период, усталость и болезни в значительной степени влияют на поведение животных. Это связано с тем, что те или иные формы поведения обусловлены внутренними факторами. Так, уровень половых гормонов в крови определяет половую доминанту, а содержание питательных веществ — пищевое поведение. Врожденные и приобретенные реакции по-разному проявляются у сытого или голодного животного, усталого или больного. Например, активная оборонительная реакция более сильно выражена у самки, имеющей детенышей, а кормодобыва-тельные рефлексы с разной силой и изобретательностью проявляются у сытого или голодного животного.
В зависимости от продуктивности меняется и поведение животных: у высокопродуктивных коров время поедания корма больше, а время лежания меньше, чем у коров низкой продуктивности. В начале лактации коровы больше времени затрачивают на кормление и меньше лежат, чем в конце лактационного периода. Глубокостельные коровы стоят больше, поедают корм быстрее и лежат меньше, чем в первые месяцы стельности.
Таким образом, сформировавшееся в процессе индивидуального развития поведение не является постоянным и незыблемым, а зависит от многих экзогенных и эндогенных факторов.
Современная наука о поведении животных основывается на последних достижениях в области общей физиологии, экологии, эволюционной теории, эндокринологии, нейрофизиологии, генетики, кормления, зоотехнии, бионики и др. Только объединенными усилиями коллективов разных научных учреждений в тесном сотрудничестве с практикой современного животноводства будут решены задачи формирования стад и групп животных, наилучшим образом приспособленных к условиям кормления и содержания в крупных специализированных промышленных хозяйствах.
ПРИЛОЖЕНИЕ
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
megalektsii.ru
Факторы, влияющие на поведение
Поведение животных постоянно изменяется, формируется и проявляется в зависимости от состояния внутренней среды организма и внешних воздействий. В одних случаях это помогает приспособиться к меняющимся условиям жизни, а в других - может носить негативный характер и привести к снижению адаптации животных и заболеваниям.
Наиболее изменчивы приобретенные реакции и сформировавшиеся условные рефлексы. Инстинкты у животных наследственно закреплены и следовательно, очень прочные, но и они иногда тоже могут изменяться или временно не проявляться. Полезные для данной популяции изменения врожденного поведения могут в дальнейшем передаваться потомству. Поэтому основные формы поведения животных эволюционируют вместе с видом.
Важнейшими факторами, влияющими на поведение животных являются: наследственность, типы высшей нервной деятельности, условия окружающей среды и физиологическое состояние.
Наследственность. Наблюдения за животными-близнецами, помещенными в различные условия, выявили сходство их основных поведенческих реакций: близкая скорость образования, дифференцировки и переделки условных рефлексов, а также таких качеств, как смелость, трусость, упрямство, отношение к человеку.
Известны породные отличия в поведении животных одного вида. Например, доберман-пинчеры характеризуются повышенной возбудимостью, а кавказские овчарки - флегматичностью. У верховых лошадей преобладают сильные, подвижные типы ВНД, а у тяжеловозов - инертные.
Наследственные факторы определяют те формы поведения, которые преимущественно базируются на безусловных рефлексах (половая активность, агрессивность, драчливость или же уклонение от внутригрупповых конфликтов). Поэтому селекция животных по показателям типов ВНД, рангов животных, их продуктивности, стрессоустойчивости может быть эффективной для формирования стада молочных коров, пригодных к условиям машинного доения.
В процессе одомашнивания велся отбор животных в желательном для человека направлении, что сопровождалось и изменениями условий их жизни. Поэтому поведение домашних животных существенно отличается от их диких предков. Некоторые инстинкты у них исчезли - миграция у уток и гусей, насиживание у кур-несушек, агрессивность по отношению к человеку. Тем не менее, многие виды биологического поведения у домашних животных сохранились, они проявляются в стаде, на пастбище, в просторных загонах - установление иерархических отношений, характерные движения во время кормления, в период половой охоты и др.
Типы высшей нервной деятельности. Животные разного типа высшей нервной деятельности по-разному проявляют себя в одинаковых ситуациях. Так, животные-лидеры обычно имеют сильный уравновешенный тип, а низших социальных рангов, как правило, имеется слабый тип нервной системы.
От типа ВНД зависит быстрота выработки и прочность закрепления условных рефлексов, динамического стереотипа, устойчивость к неблагоприятным условиям содержания.
Условия окружающей среды. Поведение направлено на удовлетворение биологических потребностей животного (питание, воспроизводство, движение, игра и т.п.) и в значительной степени обусловлено изменениями окружающей среды. Например, во время смены стойлового содержания на пастбищное меняются формы пищевого, полового и социального поведения у коров.
Периодические сезонные изменения среды (освещенность, солнечная радиация, климат, состав кормов и пр.) вызывают повторяемость определенных физиологических процессов, их качественные и количественные особенности. Эти изменения называются сезонными нишами. Примером может служить сезонный ритм воспроизводительных функций. У животных очень четко проявляются и суточные (циркадные) ритмы, связанные с вращением Земли и продолжительностью светового дня (чередование сна и бодрствования, подвижность, колебания вегетативных процессов - артериального давления, температуры тела, концентрации гормонов в крови и многое другое). Формирование общего суточного ритма поведения животных зависит от установленного распорядка дня, в частности от кратности кормления и доения.
Знание периодичности сезонного и суточного поведения животных используется в практике животноводства для повышения продуктивности. Так, искусственное увеличение продолжительности светового дня в птичнике продлевает цикл яйцекладки у кур.
Одной из причин, нарушающих поведение животных, является шум от работающих в помещении механизмов. При уровне шума 40-60 дБ животные, особенно лошади и коровы, проявляют беспокойство, у них понижаются аппетит и поедаемость кормов, сокращается время отдыха. Но к такому уровню шума они привыкают, и биологическое поведение восстанавливается. Однако при возрастании шума до 60-70 дБ продуктивность снова падает, а привыкание не возникает.
Изменяется поведение животных и при перегруппировках. При вводе в группу новичка вначале с ним «выясняют отношения» животные высших рангов, а остальные выжидают. Чужака окружают, вступают с ним в единоборство и стараются изгнать из группы, особенно если это животное высокого ранга. При введении в группу животного низкого ранга реакция группы более слабая и непродолжительная. Вводимые животные также по-разному реагируют на новую обстановку: низкого ранга сразу признают свое поражение, а высшего принимают вызов, при этом частота дыхания у них доходит до 100 при пульсе более 100 ударов в 1 мин. Отделение от группы также часто вызывает стрессовое состояние. Животное, оставленное в загоне, когда остальных уводят, проявляет сильное беспокойство и стремится соединиться с группой. Известно, что разлученные коровы или лошади, ранее симпатизировавшие друг другу, нередко похожи на больных животных: у них пропадает аппетит, отмечаются вялость, депрессия, угнетенное состояние.
При резких изменениях питания, климата, продолжительности светового дня поведение животных меняется с целью наилучшего приспособления организма к новым условиям. Так, завезенные из Канады в Сибирь бобры успели за лето построить и оборудовать жилище и запасти на предстоящую зиму корм - ветки и стволы деревьев. Однако сибирская зима более длительная и суровая, чем на их родине, и в первой зимовке им не хватило запасенного корма, но на следующий год семья бобров заготовила значительно больше корма и прекрасно перезимовала.
Сельскохозяйственные животные в условиях круглогодового стойлового содержания находятся среди относительно постоянных факторов окружающей среды (температура и влажность помещения, освещенность, гиподинамия, состав корма, одни и те же окружающие животные) и приспосабливаются к ним соответствующим образом, т.е. у них вырабатывается определенный стереотип поведения. Однако за адаптацию к слишком постоянным условиям жизни организм расплачивается потерей пластичности и способности успешно противодействовать неблагоприятным факторам. Поэтому периодическое изменение микроклимата, светового дня, а также прогулки, пастьба, игры оказывают тренирующее воздействие на многие системы организма, что способствует лучшему физиологическому развитию и разнообразию жизненных проявлений, повышению резистентности к болезнетворным факторам. В то же время частые и интенсивные вмешательства в естественный жизненный ритм животного могут вызвать неблагоприятные физиологические реакции, нарушение адаптации и стрессовое состояние.
Физиологическое состояние животных. Голод, сытость, беременность, лактация, послеродовой период, усталость и болезни в значительной степени влияют на поведение животных. Это связано с тем, что те или иные формы поведения обусловлены внутренними факторами. Так, уровень половых гормонов в крови определяет половую доминанту, а содержание питательных веществ влияет на пищевое поведение. Врожденные и приобретенные реакции по-разному проявляются у сытого или голодного животного, здорового или больного. Например, активная оборонительная реакция наиболее выражена у самки, имеющей детенышей, а кормодобывательные рефлексы с наибольшей силой и изобретательностью проявляются у голодного животного.
В зависимости от продуктивности меняется и поведение животных: у высокопродуктивных коров время поедания корма больше, а время лежания меньше, чем у коров низкой продуктивности. В начале лактации коровы больше времени затрачивают на кормление и меньше лежат, чем в конце лактационного периода. Глубокостельные коровы стоят больше, поедают корм быстрее и лежат меньше, чем в первые месяцы стельности.
Таким образом, сформировавшееся в процессе индивидуального развития поведение не является постоянным и незыблемым, а зависит от многих экзогенных и эндогенных факторов.
studfiles.net
Внешние факторы инстинктивного поведения
Количество просмотров публикации Внешние факторы инстинктивного поведения - 210
Животное способно незамедлительно и с максимальной пользой для себя реагировать на изменение в окружающей среде. Это обеспечивается тем, что соответствующие эндогенные системы периодически активируются как собственной ритмикой, так и внешними воздействиями (к примеру, изменением продолжительности светового дня, повышением или уменьшением температуры и т. д.). При этом инстинктивные движения, по этологической концепции, заблокированы специальной системой ʼʼврожденных пусковых механизмовʼʼ. Οʜᴎ представляют из себясовокупность нейросенсорных систем, обеспечивающих приуроченность поведенческих актов к биологически адекватным условиям среды (к ʼʼпусковой ситуацииʼʼ). Как только животное оказывается в такой ситуации, соответствующий врожденный пусковой механизм обеспечивает распознавание, оценку и интеграцию специфических для данной инстинктивной реакции раздражителей, после чего наступает растормаживание, снятие ʼʼблокировкиʼʼ. Очевидно, одновременно происходит активация соответствующих нервных центров и снижение порогов их раздражимости.
Характерной особенностью для врожденных пусковых механизмов является избирательность реагирования на внешние стимулы: они отзываются только на совершенно определенные комбинации раздражителей, которые только и могут вызвать биологически целесообразную реакцию.
Благодаря врожденным пусковым механизмам внутренняя мотивация поведения получает ʼʼвыход наружуʼʼ, т. е. создается возможность без индивидуального опыта в биологически значимых ситуациях реагировать так, чтобы это способствовало сохранению особи и вида.
Подводя итог сказанному, можно сказать, что под врожденным пусковым механизмом следует понимать совокупность нейросенсорных систем, обеспечивающих адекватность поведенческих актов по отношению к ʼʼпусковой ситуацииʼʼ: настройку анализаторов на восприятие специфических раздражителей и распознавание последних, интеграцию соответствующих раздражений и растормаживание (или активацию) нервных центров, связанных с данным поведенческим актом.
Внешние раздражители, составляющие в своей совокупности пусковую ситуацию, получили название ʼʼключевых раздражителейʼʼ. Ключевые раздражители являются такими признаками компонентов среды, на которые животные реагируют независимо от индивидуального опыта врожденными, видотипичными формами поведения, точнее — определенными инстинктивными движениями. В описанном поведении курицы это будут определенные общие признаки, свойственные всем ее наземным врагам.
Кроме растормаживающих ключевых раздражителей (их называют также ʼʼпусковыми раздражителямиʼʼ) различают также настраивающие ключевые раздражители, предварительно понижающие порог раздражимости нервных центров, причастных к данным действиям животного, а также направляющие ключевые раздражители, о которых пойдет речь при обсуждении таксисов. Общим свойством всех ключевых раздражителей является то, что это специфические элементарные признаки жизненно важных компонентов среды. Ключевыми раздражителями являются простые физические или химические признаки (ʼʼпростоʼʼ форма, размер, подвижность, цвет, запах и т. д.), или их пространственные отношения (взаиморасположение деталей, относительная величина и т. д.), или же векторы. Носителями этих признаков бывают как другие животные, так и растения и объекты неживой природы. В последнем случае ключевые раздражители выполняют преимущественно направляющую функцию. Так, к примеру, немецкий этолог Ф. Вальтер показал; что у детенышей антилоп ключевым стимулом, определяющим выбор места отдыха (лежа, неподвижно) является ʼʼчто-то вертикальноеʼʼ вне зависимости от того, что конкретно это за объект.
Наилучшим образом изучены ключевые раздражители, носителями которых являются животные. Эти раздражители представляют особый интерес и потому, что являются первичными, генетически фиксированными элементами общения у животных. Так, в ставших уже классическими опытах голландского зоолога Н. Тинбергена, одного из основоположников современной этологии, с помощью макетов изучалась пищевая реакция (ʼʼпопрошайничествоʼʼ) птенцов серебристых чаек (клевание клюва родительской особи) и дроздов (вытягивание шеи и раскрытие клюва) при появлении родительской особи.
Рис. 2. Опыты Тинбергена с применением макетов головы серебристой чайки. Линиями, обозначена интенсивность реакций птенцов на макет (число клеваний)
В естественных условиях голодный птенец серебристой чайки клюет красное пятно на желтом клюве родителя, и тот в ответ отрыгивает пищу в рот птенцу. В опытах предъявлялась серия все более упрощаемых моделей—макетов. Первая модель точно воспроизводила внешний облик естественного носителя ключевых раздражителей, т. е. головы взрослой серебристой чайки с желтым клювом и красным пятном на нем. В последующих моделях путем проб ʼʼпостепенно исключались отдельные детали, и в результате макет становился все менее похожим на голову птицы (рис. 2). В конце концов остался лишь плоский красный предмет с продолговатым выступом. Но данный предмет оказался способным вызвать даже более сильную реакцию птенцов, чем исходная модель. Еще более эта реакция должна быть усилена, в случае если данный макет заменить тонкой белой палочкой, исчерченной поперечными темно-красными полосами. Ключевыми раздражителями в данном примере будут просто ʼʼкрасноеʼʼ и ʼʼпродолговатоеʼʼ.
Интересные опыты проводил еще в середине 30-х годов ХХ ст. Г. А. Скребицкий совместно с Т. И. Бибиковой на подмосковном озере Киёво, изучая отношение чайки к ее яйцам. Яйца перекладывались из гнезда в гнездо, заменялись яйцами других видов, искусственными яйцами, а затем и различными посторонними предметами различной величины, формы и окраски. Оказалось, что чайки садились как на чужие, так и на деревянные, стеклянные, каменные, глиняные яйца различной величины и самой разнообразной окраски и начинали их ʼʼвысиживатьʼʼ. То же самое наблюдалось, когда вместо яиц в гнездо подкладывались разноцветные шары, камешки или картофелины. По свидетельству экспериментаторов, ʼʼчайки, сидящие на таких предметах, представляли очень оригинальную картину, но в особенности необычайным становилось зрелище, когда согнанная с гнезда птица возвращалась к нему обратно и, прежде чем сесть, заботливо поправляла клювом разноцветные шары, камешки или картошкуʼʼ.
В случае если, однако, чайкам подкладывались предметы иной формы, к примеру кубики или камни с неровными краями, поведение птиц заметно изменялось. Когда за край гнезда клалось по одному яйцу и инородному предмету, птицы вкатывали обратно в гнездо наряду с яйцами лишь округлые гладкие предметы, размеры которых соответствовали размерам яиц. В известных пределах не препятствовали этому существенные отклонения от нормы в весе (каменное яйцо весило в два с лишним раза больше, чем чаечье), материал, из которого был изготовлен предмет, и окраска. Исследователи пришли к выводу, что положительная реакция чайки на яйцо определяется лишь несколькими его элементарными признаками — округлостью, отсутствием выступов, углублений или насечек. Именно эти признаки и выступали здесь как ключевые раздражители.
Важная особенность действия ключевых раздражителей состоит по сути в том, что они подчиняются закону суммации: с увеличением их параметров пропорционально усиливается инстинктивная реакция животного. В экспериментальных условиях это может привести к так называемым ʼʼсупероптимальнымʼʼ реакциям, когда животное ʼʼпреувеличенноʼʼ, сильнее, чем в норме, реагирует на искусственный раздражитель, в котором ʼʼсгущены краскиʼʼ. Мы уже встречались с этим явлением при описании опыта с птенцами чайки, когда они сильнее реагировали на поперечно исчерченную красными полосками палочку, чем на настоящий клюв живой птицы. Суммация проистекает здесь из многократности красных меток и их большей контрастности.
Эффект супероптимальной реакции может в условиях эксперимента привести даже к биологически абсурдному поведению животного. В случае если, к примеру, предложить чайке два яйца разной величины, она закатит в гнездо более крупное. В итоге может возникнуть такое положение, что птица бросит свое яйцо, чтобы попытаться высиживать деревянный макет яйца гигантских размеров, обладающих супероптимальными признаками ключевого раздражителя.
Как видно из приведенных примеров, ключевые стимулы действуют на поведение животного принудительно, заставляя его выполнять определенные инстинктивные движения, невзирая на возможно воспринимаемую животным общую ситуацию. Это объясняет многие, ранее казавшиеся загадочными моменты в поведении животных.
В процессе эволюции возникают адаптации к более постоянным компонентам внешней среды, необходимым для удовлетворения потребностей, непрестанно возникающих в результате изменений внутренней среды организма. Нахождение (или избегание) важных для организма компонентов внешней среды осуществляется путем ориентации по типичным признакам этих компонентов—ключевым раздражителям.
Результаты этой ориентации реализуются нейросенсорными системами (врожденными пусковыми механизмами), которые действуют рефлекторно и включают эндогенные, генетически фиксированные компоненты инстинктивного поведения. Τᴀᴋᴎᴍ ᴏϬᴩᴀᴈᴏᴍ, действуя ʼʼвовнеʼʼ, врожденные пусковые механизмы обеспечивают избирательную направленность внешней активности организма лишь на определенные сигнальные стимулы; действуя же ʼʼвовнутрьʼʼ, они осуществляют оценку и отбор поступающей через рецепторы информации и ее реализацию для активации или понижения порогов раздражимости соответствующих нервных структур, для снятия ʼʼблокировкиʼʼ, растормаживания эндогенных нервных процессов, мотивирующих инстинктивные движения и действия. Τᴀᴋᴎᴍ ᴏϬᴩᴀᴈᴏᴍ, осуществляется на врожденной базе корреляция внутренних потребностей организма с биологически существенными изменениями в окружающей его среде.
Только в этой корреляции состоит все биологическое значение эндогенной мотивации поведения. Внутренние стимулы служат лишь для осуществления движения по отношению к среде, без которого организм нежизнеспособен.
Факторы поведения сами зависят от процессов обмена веществ. Эти процессы уже непосредственно связаны с окружающей организм средой. А так как организм активно регулирует, создает необходимые внешние предпосылки для нормального протекания обменных процессов как раз посредством поведения, то круг смыкается. И в данном отношении проявляется относительность независимости эндогенной спонтанной активности нервной системы, способности к ʼʼсамопрограммированиюʼʼ. Все эти эндогенные процессы лишь постольку независимы от внешней среды, поскольку они лишь опосредованно связаны с ней.
referatwork.ru
Внутренние и внешние факторы. Структура инстинктивного поведения — Мегаобучалка
Внутренние факторы инстинктивного поведения.На протяжении длительного времени считалось, что научение определяется внешними факторами, а инстинктивное поведение – исключительно внутренними факторами, причем природа этих факторов была неизвестна. Поиск, уточнение внутренних факторов инстинктивного поведения позволили бы ответить на вопрос, что же определяет мотивацию поведения.
Внутренние факторы, бесспорно, оказывают влияние на инстинктивное поведение животных. В середине XX в. американский биолог П. Уит проводил опыты с пауками, в ходе которых изучал плетение паутины при попадании в организм животного разных химических веществ. Нужное вещество наносилось в виде капли непосредственно на паутину или вводилось при помощи шприца в попавшуюся жертву. Каждое вещество стимулировало паука плести паутину определенного вида, сама же реакция плетения паутины является у паука наследственной. Так, кофеин заставлял пауков плести бесформенную паутину из беспорядочно спутанных нитей, паук при этом испытывал подобие невроза. При поступлении в организм первитина паук становился очень беспокойным и не сплетал паутину целиком. Хлоргидрат вызывал у паука оцепенение, и он не доплетал паутину. А лизергиновая кислота способствовала повышению концентрации внимания на плетении, и паук сплетал паутину очень тщательно и ровно, при этом ее качество превосходило естественное.
Внутренняя среда организма постоянна, разнообразные процессы регуляции направлены на поддержание физико-химического состава среды. Она постоянно обновляется, однако все ее параметры поддерживаются на определенном уровне за счет саморегуляции, что обеспечивает протекание всех биохимических реакций. Особенность внутренних процессов животного организма состоит в том, что они часто протекают в форме ритмов. В 1930-е гг. советский зоопсихолог В.М. Боровский выдвинул предположение о том, что именно отклонения этих внутренних ритмов организма от нормы являются первичной мотивацией поведенческих реакций. В определенных условиях внутренняя согласованность физиологических ритмов нарушается, и прежнее равновесие в новых условиях не обеспечивает нормальное функционирование организма. Возникает внутреннее побуждение, направленное на восстановление внутреннего равновесия, т. е. появляется потребность. Инстинктивное поведение в этом случае будет направлено на удовлетворение этой потребности.
Важнейшими источниками внутренних стимулов инстинктивного поведения являются гормоны и рецепторы. Известно, что половые гормоны и гормоны гипофиза стимулируют целый ряд форм поведения, связанных с размножением, – драки между самцами за самку и за территорию, охрана гнезда, брачные игры.
Для внутренней мотивации большое значение имеют прежде всего ритмические процессы, которые происходят в центральной нервной системе. Ритмическая активность ее стволовой части у позвоночных животных и брюшных нервных структур у беспозвоночных обеспечивает ориентацию поведения во времени. Известно существование у животных так называемых «биологических часов» – автономных колебательных процессов, которые осуществляют регуляцию всех ритмов жизнедеятельности организма. «Биологические часы» определяют колебания внешней активности поведения животных, все действия, которые повторяются с определенной цикличностью. Они как бы закладывают основу инстинктивного поведения животного, а факторы среды вносят в эти ритмы свои коррективы. Изменения могут быть связаны с действием разнообразных внешних стимулов (слуховых, зрительных и др.), а также могут зависеть от общего физиологического состояния животного в данный момент. Чаще всего в поведении животных отмечаются циркадные, или суточные, ритмы, период которых равен суткам.
Интересно отметить, что активность животного подвержена таким ритмическим суточным колебаниям даже в условиях полной изоляции от всех факторов естественной среды. Например, животное можно поместить в условия полного круглосуточного освещения и тем не менее наблюдать у него чередование периодов сна и бодрствования, близкое к естественным. Кроме того, в течение дня в поведении животных могут быть отмечены и краткосрочные ритмы. Примером могут послужить наблюдения немецкого этолога В. Шлейдта за индюками. Он отметил, что клохтанье индюка в течение дня повторяется с определенным ритмом, который сохраняется даже при полной изоляции птицы и лишении ее слуха.
Помимо ориентации поведения животного во времени, «биологические часы» ориентируют его в пространстве. Например, перелетные птицы при ориентации по положению солнца должны в каждый момент времени соотносить его положение со временем дня. Это происходит, когда они соотносят информацию о положении солнца с внутренними циркадными ритмами.
Внутренние факторы создают в организме состояние, предшествующее проявлению той или иной инстинктивной реакции. Однако начало этой реакции может зависеть от внешних условий среды. Например, определенный уровень половых гормонов и гормонов гипофиза стимулирует различные поведенческие реакции животного, связанные с размножением, но продуцирование этих гормонов приурочено к определенному времени года. Если животное, которое обитает в умеренной зоне Северного полушария, с наступлением весны содержать в условиях короткого светового дня, активность желез проявляться не будет. Напротив, если зимой создать животному условия постепенно увеличивающегося дня, гормоны начнут выделяться, и половое поведение проявится в зимнее время года.
Внутренние факторы обеспечивают готовность организма выполнить то или иное инстинктивное движение, внешние стимулы могут быть не обязательны для проявления инстинктивной реакции.
Немецкий нейрофизиолог Э. Хольст обнаружил в стволе головного мозга курицы несколько зон. При воздействии на эти зоны слабым электрическим током возникают инстинктивные движения, соответствующие той или иной зоне. Было отмечено, что если длительное время воздействовать на одну и ту же зону, усиливая силу раздражения, можно наблюдать целый ряд инстинктивных действий, которые будут совершаться в том же порядке, что и в естественных условиях. Например, курица выполняла движения, которые совершаются при приближении наземного хищника: проявляла легкое беспокойство, потом поднималась, хлопала крыльями, вскрикивала, а затем взлетала. При этом самого раздражителя (хищника) в пределах ее видимости не было. Таким образом, под влиянием исключительно внутренних факторов могут проявиться не только отдельные инстинктивные движения, но и целые инстинктивные действия. Однако при этом не следует забывать, что в естественных условиях инстинктивные действия «запускаются» все же внешними факторами. Приближение наземного хищника, которого увидела бы курица, привело бы к возбуждению соответствующей зоны головного мозга птицы, которая в условиях опыта была простимулирована искусственно.
Внешние факторы инстинктивного поведения.Если задача внутренних факторов инстинктивного поведения состоит преимущественно в том, чтобы подготовить организм к выполнению определенного поведенческого акта, то внешние факторы чаще играют роль своеобразных активаторов этого инстинктивного действия.
Все инстинктивные действия заблокированы специальной системой, которая получила название «врожденный пусковой механизм». Это определенная совокупность нейросенсорных систем, обеспечивающих приуроченность поведенческих инстинктивных актов к ситуации, в которой такое поведение будет наиболее биологически адекватным, т. е. к так называемой «пусковой ситуации». Врожденный пусковой механизм реагирует на те или иные внешние раздражители или на их комбинации, для него характерна высокая избирательность. Каждый раздражитель, сигнал (или их комбинация) будут специфичны для определенной инстинктивной реакции. Врожденный пусковой механизм распознает их, проводит анализ, интегрирует информацию и снимает блокировку с данной реакции. При этом порог раздражимости соответствующих нервных центров снижается, и они активируются. Внутренняя мотивация «находит выход», и инстинктивная реакция осуществляется именно в тех условиях и в той ситуации, когда она биологически значима. Австрийский этолог К. Лоренц (1903–1989) назвал такой механизм «разблокирования» инстинктивной реакции врожденной схемой реагирования.
Инстинктивное действие проявляется в ответ на свой набор внешних раздражителей. Эти раздражители получили название «ключевые», или «знаковые». Внешний сигнал в данном случае соотносят с ключом, который идеально подходит к замку (врожденному пусковому механизму). Например, в период размножения для самцов птиц ключевыми будут стимулы, характерные для самок того же вида, эти стимулы будут вызывать у самцов инстинктивные действия, связанные с ухаживанием, спариванием и т. д.
В качестве ключевых раздражителей могут выступать простые физические или химические признаки, их пространственные отношения (например, соотнесение размеров) или векторы.
Носителями ключевых раздражителей могут быть не только другие особи, но и растения, а также различные объекты неживой природы. Немецкий этолог Ф. Вальтер отметил, что у детенышей антилоп ключевым стимулом, определяющим выбор места отдыха, является любой вертикальный объект. Ключевой раздражитель выполняет здесь направляющую функцию.
Знаковые стимулы чрезвычайно разнообразны и по своей природе: они могут быть зрительными, акустическими, химическими и др. Например, в половом поведении многих насекомых, амфибий и некоторых млекопитающих ключевыми раздражителями служат химические вещества (половые аттрактанты, феромоны). К звуковым стимулам относятся разнообразные крики, песни, специфичные для определенного вида животных. Зрительные ключевые стимулы получили название «релизеры». К ним относятся различные морфологические признаки (особенности окраски тела, гребни, хохолки у птиц, наросты). Например, для самок крякв релизерами являются «зеркальца» на маховых перьях селезней. Существуют и особые видоспецифические комплексы движений, которые могут выступать в качестве знаковых стимулов (позы подчинения, угрожающие позы, приветственные ритуалы, брачные ритуалы).
Животное способно распознать ключевой раздражитель даже при первом предъявлении. Например, красное пятно на клюве чайки вызывает реакцию «попрошайничества» у птенцов. Для пояснения принципа действия этого раздражителя часто пользуются аналогией с ключом и замком.
Различают также настраивающие ключевые раздражители. Их действие отличается от знаковых раздражителей. Эти стимулы понижают порог раздражимости нервных центров и направляют ключевые раздражители.
Существование ключевых раздражителей и их роль в развитии инстинктивных реакций доказаны множеством наблюдений и опытов. Н. Тинберген изучал пищевую реакцию птенцов серебристых чаек и дроздов при появлении родительской особи методом макетов.
Естественная реакция голодного птенца чайки на родительскую особь заключается в том, что он клюет красное пятно на клюве (желтом) взрослой птицы. Тинберген в своих опытах использовал несколько макетов. Лишь один макет в точности повторял внешний облик головы взрослой серебристой чайки. На остальных макетах исключались отдельные детали, и постепенно макет становился все менее похожим на голову чайки. Последний макет представлял собой плоский красный предмет с продолговатым выступом. Однако реакция птенцов на этот предмет не только не была слабее реакции на первый макет, но даже превосходила ее. Еще более интенсивной становилась реакция птенца на макет в виде тонкой белой палочки с поперечными темно-красными полосами. Отсюда можно сделать вывод, что ключевыми раздражителями для появления реакции «попрошайничества» у птенцов серебристой чайки служат красный цвет и продолговатая форма.
В опытах с десятидневными птенцами дрозда в качестве макетов использовались плоские диски. Если птенцам дрозда предлагали круг, они тянулись к его верхней части, где предполагалась голова родительской птицы. Если к большому кругу прикрепляли маленький, птенцы начинали тянуться к нему, а когда прикрепляли два маленьких круга разного размера, решающей становилась относительная величина кругов. При большом размере «туловища» птенцы тянулись к дополнительному кругу больших размеров, при маленьком – к меньшему. Таким образом, ключевые раздражители в данном случае – взаимное расположение и относительная величина деталей макета.
Опыты по изучению ключевых стимулов у птиц проводили российские орнитологи Г.Л. Скребицкий и Т.И. Бибикова. В ходе опытов исследовалось отношение чайки к ее яйцам. Исследователи перекладывали яйца из одного гнезда в другое, заменяли их яйцами других видов птиц, иными предметами разной формы, величины, окраски. Чайки охотно начинали «высиживать» чужие яйца, а также яйца других птиц, различно окрашенные муляжи из разных материалов (стекло, глина и т. д.), посторонние предметы (шарики, картофелины, камни). Птицы не отказывались вкатывать в гнездо даже тяжелые каменные шарики, т. е. эту реакцию определял не вес «яйца». Г.Л. Скребицкий писал: «...чайки, сидящие на таких предметах, представляли очень оригинальную картину, но в особенности необычайным становилось зрелище, когда согнанная с гнезда птица возвращалась к нему обратно и, прежде чем сесть, заботливо поправляла клювом разноцветные шары, камешки или картошку».[8]
Птицы отказывались насиживать предметы, не имеющие округлой формы, например камни с острыми выступами или кубики. Ученые сделали вывод, что ключевыми стимулами для чайки были округлость предмета, отсутствие на нем выступов и углублений.
Если чайке предлагали два яйца разной величины, она начинала закатывать в гнездо более крупное. Исследователи наблюдали даже такую ситуацию, когда чайка пыталась насиживать деревянный макет яйца таких гигантских размеров, что она с трудом забиралась на него. В этой ситуации имеет место супероптимальная реакция. Животное сталкивается со сверхстимулом, который обладает супероптимальными признаками ключевого раздражителя, и начинает проявлять реакцию на него сильнее, чем в норме. Таким образом, ключевые раздражители подчинены закону суммации: с увеличением параметров раздражителя инстинктивная реакция увеличивается пропорционально. Это явление может служить объяснением повышенной реакции птенцов серебристой чайки на поперечно исчерченную красными полосками палочку.
На количественную сторону действия знаковых стимулов обратил внимание Н. Тинберген при изучении реакции преследования самки у самцов бабочки-бархатницы в период размножения. Наблюдения показали, что самец взлетает не только при приближении особей своего вида, но и при виде других летающих насекомых, а также небольших птиц и даже падающих с деревьев листьев. Ученый сделал вывод, что для бархатницы в этой ситуации первостепенное значение имеют какие-то зрительные ключевые раздражители. Химические стимулы в данном случае не могут быть знаковыми, потому что направление полета самцов никак не связано с направлением ветра, а значит, запахами они не руководствуются. Тинберген и его помощники изготовили макеты бабочек из бумаги и укрепили их на тонкой леске, привязанной к длинной удочке. У каждой серии макетов оставляли только один характерный внешний признак: цвет, размер, определенную форму. При подергивании удилища макетная бабочка начинала двигаться, что вызывало у самцов бархатницы ответную реакцию преследования. Интенсивность реакции фиксировалась наблюдением.
Результаты эксперимента показали, что реакцию преследования вызывали макеты всех цветов, но наиболее активно самцы следовали за черными «бабочками» – реакция на них была даже ярче выражена, чем при виде макетов коричневого цвета, который соответствовал естественной окраске самки бархатницы. В этом случае следует говорить об усилении зрительного стимула – темного цвета.
Аналогичная картина была получена при сравнении интенсивности реакции на размер макета. Самцы наиболее активно преследовали макеты большего размера, чем естественная величина самки. Такой стимул, как форма тела объекта, оказался для бархатниц не столь важен. Самцы реагировали на макеты всех форм, при этом наименее эффективны были длинные прямоугольные макеты. Однако наблюдения показали, что это связано с нарушением характера движения таких «бабочек»: он становился менее «танцующим».
Тинберген обратил внимание и на другую особенность действия ключевых стимулов, которую он назвал сумматором стимулирования. Ученый писал: «...слабо привлекающая белая модель вызовет тот же процент реакций, что и черная, если ее показывать на меньшем расстоянии, чем черную. Эффективность белой маленькой модели также заметно усиливается, если заставить ее „танцевать“. Таким образом, недостаточную эффективность одного параметра можно скомпенсировать усиленной привлекательностью совершенно иного параметра... стимулы складываются в некоем „сумматоре стимулирования“, который и заставляет бархатницу реагировать соответствующим образом».[9]
Кроме того, Тинберген отметил, что состояние самца определяет, какие стимулы в данный момент входят в этот сумматор. Например, в обычных условиях самцы реагировали только на тон окраски макета (темный или светлый), т. е. сами цвета в сумматор не входили. При кормлении же на макетах, окрашенных в яркий цвет, у самцов отмечалась реакция исключительно на голубые и желтые модели, т. е. знаковым стимулом становился цвет.
Реакция на ключевой стимул не всегда бывает адекватна ситуации и может не привести к желаемому результату. Так, Тинберген описывает явление, получившее название «осечки». Осечка в поведении животного происходит при столкновении со «сверхстимулом». Примером такого «сбоя» может служить выкармливание певчими птицами птенца кукушки. Ключевыми стимулами, заставляющими птицу-родителя кормить птенца, являются большой клюв и ярко окрашенный зев птенца. Оба эти признака у кукушонка имеют «сверхнормальное» выражение. Тинберген пишет: «Вполне возможно, что многие певчие птицы не только кормят птенца кукушонка, но и получают удовольствие из-за его огромного и привлекательного рта».[10]
Осечка может происходить и во взаимоотношениях представителей разных классов животных. Описан случай, когда птица кардинал на протяжении нескольких недель кормила насекомыми золотых рыбок в бассейне. Птица реагировала на широко открытый рот рыбок, являющийся для нее ключевым стимулом при выкармливании птенцов.
В заключение следует отметить, что инстинктивное поведение животных чаще всего обусловлено не отдельными факторами, а их комплексом. При этом необходимо сочетание внешних и внутренних факторов. Например, голуби кормят своих птенцов, отрыгивая «зобное молочко», богатое протеинами. Сам процесс образования в зобе «молочка» стимулируется выделением гормона пролактина (внутренний стимул). Однако реакция отрыгивания вызывается не наполнением зоба, а внешней стимуляцией со стороны птенца, который своей тяжестью давит на зоб родителя. В зимнее время даже при наполненном пищей зобе у голубя не происходит такой реакции, потому что нет внешней стимуляции.
Структура инстинктивного поведения.Еще в начале XX в. американский исследователь У. Крэг («Влечения и антипатии как составляющие инстинкта», 1918) показал, что любое инстинктивное действие состоит из отдельных фаз. Крэг выделил две фазы, которые получили названия: поисковая (подготовительная) фаза, или аппетентное поведение, и завершающая фаза (завершающий акт).
Крэг показал, что в естественных условиях животные ищут те ключевые стимулы или их комбинации (пусковые ситуации), которые необходимы для осуществления определенной инстинктивной реакции. Например, животные ищут пищу, особей другого пола в период размножения, места для гнездования и т. д. Крэг назвал эти поисковые формы поведения аппетентными, а состояние животного в этот момент – аппетенцией. Воспринимаемые на поисковой фазе поведения промежуточные раздражители не являются для животного целью, они необходимы лишь для того, чтобы привести к восприятию ключевых стимулов завершающего поведения. Завершающая фаза инстинктивного поведения представляет собой само потребление животным необходимых ему элементов среды, именно эта фаза и является непосредственно инстинктивным поведением.
Завершающая фаза наследственно определена, видотипична, в ней заключается биологический смысл всего инстинктивного действия. Эта фаза поведения состоит из небольшого количества движений, всегда выполняемых в четкой последовательности. Она стереотипна, определяется строением тела животного. На этой фазе возможны лишь незначительные индивидуальные вариации поведения, которые определяются генетически. Приобретенные компоненты поведения практически не играют роли в завершающем акте, а чаще всего и вовсе отсутствуют. К. Лоренц назвал завершающие акты поведенческих реакций эндогенными движениями, они видотипичны, наследственны и не требуют специального обучения.
В отличие от завершающего акта поисковая фаза более изменчива и адаптивна по отношению к условиям, хотя также является типичной для вида. В ней переплетаются врожденные и приобретенные формы поведения, индивидуальный опыт животного. Для поискового поведения характерна ориентировочно-исследовательская деятельность животного. Именно за счет изменений аппетентного поведения инстинктивные реакции могут быть пластичными. Подготовительная фаза всегда делится на несколько этапов. Ее окончание наступает, когда животное достигнет ситуации, в которой может осуществиться следующее звено данной цепи реакций. Например, выбор самцом гнездовой территории иногда требует только возвращения на старую, прошлогоднюю территорию, а подчас может потребовать длительных поисков и даже борьбы с другими самцами. По представлению К. Лоренца, поисковую фазу поведенческого акта следует относить к целенаправленному поведению. На этой стадии совершаются различные действия, но все они подчинены определенной цели. Поисковая фаза очень важна и является для животного такой же первостепенной жизненной необходимостью, как и потребление на завершающей фазе. Именно аппетентное поведение является средством индивидуального приспособления животных к меняющейся окружающей среде. К этой фазе поведенческого акта относятся проявления элементарной рассудочной деятельности животных. Для достижения определенной конечной цели животное выбирает какой-либо путь, при этом оно оперирует понятиями и законами, которые связывают предметы и явления внешнего мира.
Концепцию о двух фазах инстинктивного поведения Крэг построил на данных, полученных в результате изучения пищевого поведения животных. Хищник, испытывающий ощущение голода, начинает искать добычу. Однако вначале он не имеет сведений о ее местонахождении и поэтому его поисковая деятельность пока ненаправленна. Вскоре хищник видит потенциальную жертву, от которой исходит первый ключевой стимул, например размер и детали окраски, и его поисковое поведение переходит на следующую стадию, которая уже имеет определенное направление. Хищник начинает уточнять местоположение, скорость передвижения добычи, ориентируясь при этом и на другие ключевые стимулы. Затем хищник преследует добычу или незаметно подкрадывается к ней, после чего схватывает и умерщвляет. Если в этом есть необходимость, жертва перетаскивается на другое место, где разделывается на куски. Только после этого поведение животного вступает в завершающую фазу, которая включает в себя непосредственное поедание добычи. Все действия животного, связанные с поиском, ловлей и умерщвлением жертвы, относятся к аппетентному поведению. Все они имеют под собой инстинктивную основу, но в большой степени зависят от процесса индивидуального научения, опыта животного и ситуации.
На каждом этапе поискового поведения есть свои подготовительные и завершающие фазы. Окончание одного этапа является сигналом к началу следующего и т. д. Последовательные этапы часто имеют несколько степеней соподчинения, поэтому складывается сложная структура поведения животного. Например, поисковое поведение может привести не к завершающей фазе поведенческого акта, а к сочетанию раздражителей, стимулирующему следующую фазу поискового поведения. Примером может послужить поисковое поведение птиц в период размножения. Вначале происходит выбор территории для гнезда. Когда она найдена, начинается следующая стадия поискового поведения – строительство гнезда, затем следующая – ухаживание за самкой и т. д.
Поведение животного в значительной степени состоит из циклов, которые, в свою очередь, складываются из серии повторяющихся простых актов. Например, птица, занятая постройкой гнезда, совершает это по определенной схеме. Вначале она отправляется на поиски строительного материала, затем, отыскав его, оценивает пригодность. Если материал устраивает птицу, она несет его к гнезду, в противном случае бросает и ищет новый. Прилетев в гнездо, птица определенными движениями вплетает принесенные материалы в его структуру, формирует форму гнезда, а затем вновь улетает на поиски. Этот цикл начинается спонтанно и продолжается до тех пор, пока у птицы есть потребность в достраивании гнезда. Переключение на каждую следующую стадию поведенческой реакции осуществляется при восприятии определенного внешнего стимула. Н. Тинберген приводит пример с самками перепончатокрылых насекомых – филантов (пчелиных волков), которые выкармливают своих личинок медоносными пчелами. Оса, чтобы сделать запасы, летит в места скопления пчел, где беспорядочно летает до тех пор, пока не встретит подходящую жертву. Заметив летящее насекомое, оса подлетает к нему с подветренной стороны и останавливается примерно в 70 см. Если после этого оса уловит запах пчелы, который и будет ключевым раздражителем для перехода на следующую стадию поведенческой реакции, она схватит пчелу. Если пчелу лишить запаха с помощью эфира, оса не станет хватать ее. Следующей стадией поведения осы будет парализация жертвы ударом жала. Для начала этой стадии необходим стимул, связанный с прикосновением кжертве. Если предъявить осе макет пчелы, который на ощупь не похож на нее, но имеет тот же запах, оса не станет жалить такой макет. Таким образом, при прохождении животным различных стадий поведенческой реакции происходит смена стимулов, которые являются для него ключевыми в данный момент.
Состояние аппетенции происходит в условиях очень высокой возбудимости нервных центров, координирующих определенные физиологические реакции. К. Лоренц ввел понятие «специфический потенциал (энергия) действия». Этот потенциал накапливается под действием ряда внешних (температура, освещенность) и внутренних факторов (гормоны) в нервных центрах. Превысив определенный уровень, накопленная энергия высвобождается, после чего начинается поисковая фаза поведенческого акта. При усиленном накоплении «специфической энергии действия» завершающий акт может осуществиться спонтанно, т. е. в отсутствие соответствующих раздражителей, это так называемая «реакция вхолостую».
Для объяснения нейрофизиологического механизма этих явлений Лоренц предложил свою теорию. Базой для этой теории послужили данные немецкого физиолога Э. Хольста.
Хольст в своих опытах сосредоточил внимание на ритмической активности центральной нервной системы. Он отметил, что в изолированной брюшной нервной цепочке земляного червя можно наблюдать ритмические разряды импульсов, которые точно соответствуют сокращению сегментов червя. В дальнейших исследованиях Хольст изучал механизм плавания угря. Он фиксировал средние сегменты его тела и не давал им сокращаться. Согласно рефлекторной теории в таком случае задние сегменты тела не получат раздражения, а следовательно, также не смогут сокращаться. Однако на самом деле они приходят в движение после определенного промежутка времени. Если перерезать дорсальные корешки спинного мозга угря, тем самым нарушив передачу сенсорной информации, угорь сохранит способность к плавательным движениям, причем координация их не нарушится. Таким образом, движения тела угря выполняются не по механизму рефлекторной дуги (в зависимости от внешних раздражений), а подчиняясь ритмическим разрядам импульсов в центральной нервной системе. Опыты других ученых подтвердили это. Например, было отмечено, что удецеребрированных (с удаленными большими полушариями) кошек могут ритмически сокращаться мышцы-антагонисты ноги, полностью лишенные чувствительных нервов. Головастики и рыбы с одним неразрушенным чувствительным нервом сохраняют способность к плаванию, нормальной координации движений. Это означает, что центральной нервной системе присущ эндогенный автоматизм, который не зависит от внешних стимулов. При этом необходим минимальный уровень афферентной импульсации для поддержания возбуждения, («специфической энергии действия») в соответствующих нервных центрах на определенном уровне.
Работы Э. Хольста и его коллег подтвердили, что уровень возбуждения в соответствующих нервных центрах оказывает влияние на характер протекания инстинктивных реакций. Опыты проводились на курах, которым производили раздражение током стволовых структур мозга. В зависимости от локализации раздражаемой структуры исследователи отмечали элементарные поведенческие реакции (повороты головы, клевание) или сложные акты поведения (ухаживание). И если простые реакции всегда протекали примерно одинаково, независимо от параметров раздражения и окружающих условий, то сложные поведенческие реакции зависели от этих факторов. Так, при слабой силе тока петух набрасывался на чучело хорька, а при усилении тока – даже на руку исследователя (неспецифический раздражитель).
Гидравлическая модель К. Лоренца.Лоренц предложил гипотетическую модель осуществления инстинктивных реакций. Общие принципы ее работы ученый заимствовал из гидравлики, поэтому она получила название «гидравлическая модель».
«Специфическая энергия действия» представлена в этой модели водой, которая постепенно наполняет резервуар (происходит накопление энергии) через открытый кран, обозначающий непрерывный поток потенциальной энергии, образующийся в процессе жизнедеятельности организма. Вода (энергия) попадает в резервуар (организм) до тех пор, пока организм испытывает потребность в данной форме поведения. Давление жидкости внутри резервуара постоянно повышается, создавая определенное напряжение в системе. Отток воды из резервуара, обозначающий активность животного, происходит по трубам, ему препятствует клапан (центральный затормаживающий механизм). Клапан может открыться в двух ситуациях: при большом давлении воды, которая накопилась в резервуаре, или под действием тяжести груза, подвешенного к клапану. Груз обозначает влияние внешних раздражителей, специфичных для данного поведенческого акта. Нарастающее давление воды (накопление специфической энергии действия) и тяжесть груза (сила внешних раздражителей) суммируют свое действие на клапан. Чем сильнее стимул, тем меньший уровень энергии требуется. И напротив, чем больше накопилось энергии, тем меньшая сила внешнего стимула нужна для осуществления инстинктивной реакции. Если уровень энергии очень высок, клапан может открыться и без внешнего стимула, за счет давления воды. Это и соответствует «реакции вхолостую» (по Лоренцу, «реакция в пустоте»). Так, Лоренц описал поведение голодного скворца, который в отсутствие какого-либо раздражителя из внешней среды, например насекомого, фиксирует его взглядом и «ловит» в воздухе. Наклонный лоток с отверстиями, расположенными на разном уровне, обозначает разные типы двигательной активности животного в ходе поведенческого акта. Самое нижнее отверстие соответствует двигательной активности с самым низким порогом, остальные отверстия – формам активности с более высоким порогом. Если клапан лишь слегка приоткроется, вода вытечет в небольшом количестве и попадет только в область нижнего отверстия. Если клапан откроется больше и интенсивность тока воды возрастет, она попадет и в следующие отверстия. Когда резервуар опорожняется («специфическая энергия действия» иссякает), данный поведенческий акт прекращается.
Модель Лоренца хорошо объясняет ситуацию, когда порог совершения действия снижается при долгом его невыполнении (вода накапливается в резервуаре до такого уровня, когда потребуется небольшой груз, чтобы открыть клапан), и возможность появления реакций на неспецифические раздражители (накопление воды в резервуаре до такого уровня, когда груз для открытия клапана не нужен).
Гидравлическая модель неоднократно критиковалась из-за механистичности построения и схематичности. Понятиям «специфическая энергия действия» и «ключевые стимулы» в современной зоопсихологии соответствует понятие «специфическое побуждение».
Иерархическая теория инстинкта И. Тинбергена.Отмечено, что стереотипные двигательные реакции находятся в определенной взаимосвязи. Иногда инстинктивные движения проявляются совместно, причем повышение порога одной из них вызывает повышение порога второй. Отсюда можно сделать вывод, что обе они зависят от одного функционального «центра». Отмечается некоторая регулярность в последовательности проявления действий в сложных инстинктивных реакциях. Примером могут послужить агрессивные столкновения рыб при разделе территории. У рыб семейства цихлид непосредственным столкновениям предшествует особая демонстрация запугивания. У одних видов цихлид демонстрации кратковременны, и рыбки практически сразу переходят к нападению. У других цихлид столкновения с ранениями происходят только при равных силах самцов, а демонстрации очень сложны и длительны. Есть и такие цихлиды, у которых драки совсем отсутствуют, а ритуализированная церемония запугивания противника выполняется до истощения одного из самцов, который отступает. Подобные ритуалы представляют собой последовательные действия, начинающиеся с демонстрации боков тела, потом спинные плавники поднимаются вертикально, за этим следуют удары хвостом. Соперники могут оценить силу такого удара с помощью боковой линии, которая воспринимает колебания воды. Затем противники встают друг перед другом, и у одних видов начинаются взаимные толчки с открытой пастью, а у других противники кусают друг друга в открытые рты. Ритуалы продолжаются до тех пор, пока один соперник не устает, его окраска в этом случае бледнеет и он уплывает. Все двигательные реакции во время ритуальной демонстрации строго стереотипны и четко следуют одна за другой. Так, удары хвостом не могут начаться до подъема спинного плавника, а взаимные толчки происходят лишь после ударов хвостом.
megaobuchalka.ru
Внешние факторы инстинктивного поведения, Глава 2. Инстинктивное поведение, Часть I. Общая характеристика психической деятельности животных. Основы зоопсихологии 1976. Фабри К. Э. Страница 9. Читать онлайн
Глава 2. Инстинктивное поведение.
. . .Внешние факторы инстинктивного поведения.
Когда говорят об автономности внутренних факторов поведения, об их независимости от внешней среды, то необходимо помнить, что эта независимость является лишь относительной. Уже из приведенных опытов Хольста видно, что эндогенная активность не существует ни "сама по себе", ни "сама для себя": значение этих спонтанных процессов в центральной нервной системе состоит в предиспозиции к возникновению жизненно важных ситуаций ("в случае чего - все готово"). В результате животное способно по первому же сигналу незамедлительно и с максимальной пользой для себя реагировать на изменение в окружающей среде.
Эта готовность обеспечивается тем, что соответствующие эндогенные системы периодически активируются как собственной ритмикой, так и внешними воздействиями (например, изменением продолжительности светового дня, повышением или уменьшением температуры и т. д.). Однако инстинктивные движения, по этологической концепции, заблокированы специальной системой "врожденных пусковых механизмов". Последние представляют собой совокупность нейросенсорных систем, обеспечивающих приуроченность поведенческих актов к биологически адекватным условиям среды (к "пусковой ситуации"). Как только животное оказывается в такой ситуации, соответствующий врожденный пусковой механизм обеспечивает распознавание, оценку и интеграцию специфических для данной инстинктивной реакции раздражителей, после чего наступает растормаживание, снятие "блокировки". Очевидно, одновременно происходит активация соответствующих нервных центров и снижение порогов их раздражимости.
Характерной особенностью для врожденных пусковых механизмов является избирательность реагирования на внешние стимулы: они отзываются только на совершенно, определенные комбинации раздражителей, которые только и могут вызвать биологически целесообразную реакцию. Иными словами, в сенсорной сфере существует некая "фильтрующая" функция, выражающаяся в специфической преадаптационной "готовности" воспринимать такие раздражители.
Итак, благодаря врожденным пусковым механизмам внутренняя мотивация поведения получает "выход наружу", т. е. создается возможность без индивидуального опыта в биологически значимых ситуациях реагировать так, чтобы это способствовало сохранению особи и вида.
Подводя итог сказанному, можно сказать, что под врожденным пусковым механизмом следует понимать совокупность нейросенсорных систем, обеспечивающих адекватность поведенческих актов по отношению к "пусковой ситуации": настройку анализаторов на восприятие специфических раздражителей и распознавание последних, интеграцию соответствующих раздражений и растормаживание (или активацию) нервных центров, связанных с данным поведенческим актом,
Внешние раздражители, составляющие в своей совокупности пусковую ситуацию, получили название "ключевых раздражителей", поскольку они подходят к своим врожденным пусковым механизмам, как ключ к замку. Ключевые раздражители являются такими признаками компонентов среды, на которые животные реагируют независимо от индивидуального опыта врожденными, видотипичными формами поведения, точнее - определенными инстинктивными движениями. В описанном поведении курицы это будут определенные общие признаки, свойственные всем ее наземным врагам.
Кроме собственно растормаживающих ключевых раздражителей (их называют также "пусковыми раздражителями") различают также настраивающие ключевые раздражители, предварительно понижающие порог раздражимости нервных центров, причастных к данным действиям животного, а также направляющие ключевые раздражители, о которых пойдет речь при обсуждении таксисов. Общим свойством всех ключевых раздражителей является то, что это специфические элементарные признаки жизненно важных компонентов среды. Ключевыми раздражителями являются простые физические или химические признаки ("просто" форма, размер, подвижность, цвет, запах и т. д.), или их пространственные отношения (взаиморасположение деталей, относительная величина и т. д.), или же векторы. Носителями этих признаков могут быть как другие животные, так и растения и объекты неживой природы. В последнем случае ключевые раздражители выполняют преимущественно направляющую функцию. Так, например, немецкий это-лог Ф. Вальтер показал, что у детенышей антилоп ключевым стимулом, определяющим выбор места отдыха (лежа, неподвижно) является "что-то вертикальное" вне зависимости от того, что конкретно это за объект.
Наилучшим образом изучены ключевые раздражители, носителями которых являются животные. Эти раздражители представляют особый интерес и потому, что являются первичными, генетически фиксированными элементами общения у животных. Так, в ставших уже классическими опытах голландского зоолога Н. Тинбергена, одного из основоположников современной этологии, с помощью макетов изучалась пищевая реакция ("попрошайничество") птенцов серебристых чаек (клевание клюва родительской особи) и дроздов (вытягивание шеи и раскрытие клюва) при появлении родительской особи.
Рис. 7. Опыты Тинбергена с применением макетов головы серебристой чайки. Линиями, обозначена интенсивность реакций птенцов на макет (число клеваний).В естественных условиях голодный птенец серебристой чайки клюет красное пятно на желтом клюве родителя, и тот в ответ отрыгивает пищу в рот птенцу. В опытах предъявлялась серия все более упрощаемых моделей - макетов. Первая модель точно воспроизводила внешний облик естественного носителя ключевых раздражителей, т. е. головы взрослой серебристой чайки с желтым клювом и красным пятном на нем. В последующих моделях путем проб постепенно исключались отдельные детали, и в результате макет становился все менее похожим на голову птицы (рис. 7). В конце концов остался лишь плоский красный предмет с продолговатым выступом. Но этот предмет оказался способным вызвать даже более сильную реакцию птенцов, чем исходная модель. Еще более эта реакция может быть усилена, если этот макет заменить тонкой белой палочкой, исчерченной поперечными темно-красными полосами. Ключевыми раздражителями в данном примере будут просто "красное" и "продолговатое".
В опытах, проведенных Тинбергеном с десятидневными птенцами дроздов, выявилась другая категория ключевых раздражителей - взаиморасположение и относительная величина деталей объекта-носителя. В качестве макетов использовались плоские диски. Если показать птенцам такой круг, они будут тянуться к его верхней части (местонахождению головы птицы). Если же к большому кругу прибавить маленький, птенцы потянутся к нему. Если, наконец, прибавить к большому кругу два меньших круга, но разных размеров, то решающее значение приобретет относительная величина этих фигур. Размеры "головы" не должны приближаться к размерам "туловища": при большом "туловище" птенцы потянутся к большему дополнительному кругу ("голове") (рис. 8), при маленьком же - к меньшему.
Интересные опыты проводил еще в середине 30-х годов Г. А. Скребицкий совместно с Т. И. Бибиковой на подмосковном озере Киёво, изучая отношение чайки к ее яйцам. Яйца перекладывались из гнезда в гнездо, заменялись яйцами других видов, искусственными яйцами, а затем и различными посторонними предметами различной величины, формы и окраски. Оказалось, что чайки садились как на чужие, так и на деревянные, стеклянные, каменные, глиняные яйца различной величины и самой разнообразной окраски и начинали их "высиживать". То же самое наблюдалось, когда вместо яиц в гнездо подкладывались разноцветные шары, камешки или картофелины. По свидетельству экспериментаторов, "чайки, сидящие на таких предметах, представляли очень оригинальную картину, но в особенности необычайным становилось зрелище, когда согнанная с гнезда птица возвращалась к нему обратно и, прежде чем сесть, заботливо поправляла клювом разноцветные шары, камешки или картошку" 34.
34 Г. Скребицкий. Отношение чайки к гнезду, яйцам и птенцам. - В кн.: "Рефлексы, инстинкты и навыки" (ред. В. М. Боровский). М., 1936, стр. 144-145.
Рис. 8. Опыты Тинбергена с птенцами дрозда.Если, однако, чайкам подкладывались предметы иной формы,, например кубики или камни с неровными краями, поведение птиц, заметно изменялось. Когда за край гнезда клалось по одному яйцу и инородному предмету, птицы вкатывали обратно в гнездо наряду с яйцами лишь округлые гладкие предметы, размеры которых соответствовали размерам яиц. В известных пределах не препятствовали этому существенные отклонения от нормы в весе (каменное яйцо весило в два с лишним раза больше, чем чаечье), материал, из которого был изготовлен предмет, и окраска.
Исследователи пришли к выводу, что положительная реакция чайки на яйцо определяется лишь несколькими его элементарными признаками - округлостью, отсутствием выступов, углублений или насечек (рис. 9). Именно эти признаки и выступали здесь как ключевые раздражители.
Рис. 9. Вкатывание в гнездо различных предметов в зависимости от их свойств у обыкновенной чайки (в процентах). Верхний ряд, слева направо: яйца (чаечьи, куриные, деревянные и каменные), деревянные шары, другие округлые предметы (картошка, камни), удлиненное глиняное яйцо, глиняное яйцо с углублениями, глиняный шар с зарубками. Нижний ряд, слева направо: деревянный конус, деревянный цилиндр, деревянные и глиняные многоугольники, деревянные и глиняные кубики, угловатые камни (по Скребицкому и Бибиковой).Важная особенность действия ключевых раздражителей заключается в том, что они подчиняются закону суммации: с увеличением их параметров пропорционально усиливается инстинктивная реакция животного. В экспериментальных условиях это может привести к так называемым "супероптимальным" реакциям, когда животное "преувеличенно", сильнее, чем в норме, реагирует на искусственный раздражитель, в котором "сгущены краски". Мы уже встречались с этим явлением при описании опыта с птенцами чайки, когда они сильнее реагировали на поперечно исчерченную красными полосками палочку, чем на настоящий клюв живой птицы. Суммация проистекает здесь из многократности красных меток и их большей контрастности.
Эффект супероптимальной реакции может в условиях эксперимента привести даже к биологически абсурдному поведению животного. Если, например, предложить чайке два яйца разной величины, она закатит в гнездо более крупное. В итоге может возникнуть такое положение, что птица бросит свое яйцо, чтобы попытаться высиживать деревянный макет яйца гигантских размеров, обладающих супероптимальными признаками ключевого раздражителя.
Как видно из приведенных примеров, ключевые стимулы действуют на поведение животного принудительно, заставляя его выполнять определенные инстинктивные движения, невзирая на возможно воспринимаемую животным общую ситуацию. Это объясняет многие, ранее казавшиеся загадочными моменты в поведении животных.
Так, например, еще в начале нашего века один из авторов антропоморфических сочинений по психологии животных Т. Целл дал следующий ответ на вопрос о том, почему крупные хищники в нормальных условиях при встрече с человеком не нападают на него: например, лев не нападает не потому, что почитает человека, а потому, что неуверен в исходе схватки: "Есть ли у человека оружие, да какое оно, это оружие? - думает лев. - Да ну его, пойду-ка я лучше своей дорогой".
Конечно, лев, как и другие крупные хищники, как правило, уклоняется от нападения на человека не потому, что руководствуется такими глубокомысленными рассуждениями. Разгадка "почтительного" отношения диких зверей к человеку, очевидно, кроется в следующем. Будучи сытым, хищник не реагирует и на присутствие животных, которыми обычно питается. У голодного же хищника преследование животных-жертв и нападение на них обусловливаются сочетанием рассмотренных выше внутренних факторов (первопричина - изменение уравновешенности внутренней среды организма в результате недостатка питательных веществ) с соответствующими внешними стимулами - ключевыми раздражителями, носителями которых являются естественные объекты питания, т. е. животные-жертвы, но не человек.
Даже самый кровожадный хищник не волен нападать на кого угодно и когда угодно. Эти действия также ориентируются во времени и пространстве ключевыми раздражителями, как и все прочие поведенческие акты. Другими словами, дело не в том, "хочет" или "не хочет" животное поступить так или иначе. Если внутреннее состояние животного соответствует определенной внешней пусковой ситуации, то оно волей-неволей вынуждено вести себя так, как это диктует для данных условий генетически зафиксированный код, видотипичного поведения.
* * *Итак, в процессе эволюции возникают адаптации к более постоянным компонентам внешней среды, необходимым для удовлетворения потребностей, непрестанно возникающих в результате изменений внутренней среды организма. Нахождение (или избегание) важных для организма компонентов внешней среды осуществляется путем ориентации по типичным признакам этих компонентов - ключевым раздражителям.
Результаты этой ориентации реализуются нейросенсорными, системами (врожденными пусковыми механизмами), которые действуют рефлекторно и включают эндогенные, генетически фиксированные компоненты инстинктивного поведения. Таким образом, действуя "вовне", врожденные пусковые механизмы обеспечивают избирательную направленность внешней активности организма лишь на определенные сигнальные стимулы; действуя же "вовнутрь", они осуществляют оценку и отбор поступающей через рецепторы информации и ее реализацию для активации или понижения порогов раздражимости соответствующих нервных структур, для снятия "блокировки", растормаживания эндогенных нервных процессов, мотивирующих инстинктивные движения и действия. Таким образом осуществляется на врожденной основе корреляция внутренних потребностей организма с биологически существенными изменениями в окружающей его среде.
Психология bookap
Только в этой корреляции состоит все биологическое значение эндогенной мотивации поведения. Внутренние стимулы служат лишь для осуществления движения по отношению к среде, без которого организм - именно как саморегулирующаяся система - нежизнеспособен. И в этом, широком, смысле деятельность всей нервной системы в целом всегда рефлекторна.
Равным образом даже наиболее как будто "далеко упрятанные" от внешней среды факторы поведения сами зависят от процессов обмена веществ. Эти процессы уже непосредственно связаны с окружающей организм средой. А так как организм активно регулирует, создает необходимые внешние предпосылки для нормального протекания обменных процессов как раз посредством поведения, то круг смыкается. И в этом отношении проявляется относительность независимости эндогенных автоматизмов, спонтанной активности нервной системы, способности к "самопрограммированию". Ясно, что все эти эндогенные процессы лишь постольку независимы от внешней среды, поскольку они лишь опосредованно связаны с ней.
bookap.info
Содержание:
ФГОУ ВПО « Московская государственная академия ветеринарной медицины и биотехнологии им. К. И. Скрябина»
Кафедра физиологии
Реферат на тему: « Виды поведения животных»
Москва 2007 год
1. Этология, зоопсихология. Методы этологии.
2. Физиологические механизмы функциональной системы поведения.
3. 2 типа поведения.
4. Врожденные формы поведения:
- безусловные рефлексы, их характеристика, классификация.
- инстинкты, виды и свойства инстинктов.
5. Приобретенные формы поведения. Формы научения:
- облигатное неассоциативное научение
- факультативное ассоциативное научение
- обучение по методу проб и ошибок
- вероятностное научение
- научение «инсайт»
6. Формы поведения:
- локомоция
- пищевое поведение
- гомеостатическое и комфортное поведение
- пассивное или активное оборонительное поведение
- исследовательское поведение
- игровое поведение
- подражательное поведение
- экстраполяционное поведение
- поведение в экстремальных ситуациях
- поведение в критических ситуациях
- аномальное (ненормальное) поведение
- аутопрофилактика и аутосонация как формы поведения
- сон
- половое поведение
- родительское поведение
- типы социального поведения
- стадное поведение
- ритуальные формы поведения
7. Список литературы
Этология, зоопсихология. Методы этологии.
Изучением поведения животных занимаются две науки: этология и зоопсихология.
Этология (от греч. этос- привычка, нрав, поведение илогос- учение; термин введен впервые в 1908 году Долло) как наука представляет систему достоверных знаний биологических основ, закономерностей и механизмов поведенческих актов животных. Поведенческим актом называется целенаправленная деятельность организма для удовлетворения тех или иных биологических потребностей.
Зоопсихология- раздел учения о поведении животных, изучающий организацию высшей нервной деятельности, ее сложных механизмов. Зоопсихология ищет общие законы поведения.
Этология изучает поведение животных методами наблюдения и регистрации, а механизмы поведенческих актов- физиологическими методами.
Основным методом этологических исследований является наблюдение с регистрацией двигательных актов различными способами: протоколированием с хронометражем событий, фотографированием, киносъемкой, экспериментом (широко используют в естественных и искусственных условиях).Для изучения каждой формы поведенческих реакций предназначена своя система методов и методик.
На современном этапе развития этологии происходит объединение трех направлений- этологии, зоопсихологии и физиологии- и формирование единой науки о поведении с широким использованием методов физиологии кроме перечисленных этологических.
Физиологические механизмы функциональной системы поведения.
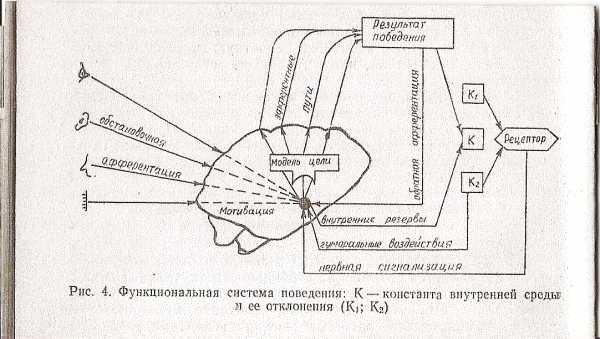
Одним из главных условий поддержания жизни является сохранение стабильности внутренней среды организма. В то же время в результате непрерывно идущего обмена веществ практически каждая константа постоянно смещается. Это заставляет организм совершать целенаправленную деятельность во внешней среде и потреблять или, наоборот, выделять в нее определенные вещества. Еще в 1916 году И. П. Павлов назвал такую деятельность «рефлексом цели», понимая под этим влечение к определенным раздражителям внешней среды, что и обеспечивает саморегуляцию организма. Он писал, что «живой организм как система существует среди окружающей природы только благодаря непрерывному уравновешиванию этой системы с внешней средой, то есть благодаря определенным реакциям живой системы на падающие на нее извне раздражения, что у более высших животных осуществляется преимущественно при помощи нервной системы в виде рефлексов».
Начиная с 1935 года П. К. Анохин развивает учение И. П. Павлова и указывает, что аппаратом саморегуляции в организме служат функциональные системы- динамические организации, главными признаками которых являются:
наличие полезного результата (системообразующего фактора)
осуществление последовательно протекающих процессов с конкретными узловыми механизмами (внутренняя архитектоника)
оценка и сличение полученного результата с внутренней моделью цели поведения.
Пусковым стимулом целенаправленного поведения является отклонение жизненно важных констант внутренней среды организма от оптимального уровня. К таким константам относятся концентрация питательных веществ и гормонов, газовый состав, осмотическое давление, температура, кислотность внутренней среды и др. Эти устойчивые отклонения жизненно важных констант от уровня, создающего комфорт и оптимальную деятельность, получили название потребностей организма.Так, снижение уровня глюкозы в крови субъективно ощущается как голод и вызывает потребность в пище. При возрастании осмотического давления возникает ощущение жажды и потребность в воде.
Отклонение константы воспринимается специальными избирательно чувствительными структурами- рецепторами (блок входа системы). Во всех тканях организма и полостях органов имеются баро-, механо-, термо-, хемо-, осморецепторы, т.е. структуры, воспринимающие давление, механические, температурные, химические воздействия и изменение осмотического давления крови или лимфы. В наиболее часто раздражаемых участках организма рецепторы расположены особенно густо (рецепторные зоны). В стенках кровеносных сосудов выделяют зоны в дуге аорты, в каротидном синусе (сонная артерия) и в месте впадения полых вен в сердце. Таким образом, контролируется кровь, выброшенная из сердца в большой круг кровообращения, направляющаяся к головному мозгу и возвращающаяся к сердцу. Специальные рецепторные зоны обнаружены в мозге, особенно в гипоталамусе и ретикулярной формации среднего мозга. Основное свойство всех рецепторов- специфичность, т.е. каждый из них воспринимает отклонение только определенной константы и не реагирует на изменение других.
Возникающие в рецепторах импульсы (пусковая афферентация) передаются по нервным волокнам к определенным центрам мозга (подкорковые нейроны) и возбуждают их. Кроме нервной сигнализации подключается еще и гуморальное воздействие на мозг (раздражающее действие определенных веществ крови), которое обеспечивает значительную продолжительность активного состояния центров.
Возникающий на основе пусковой сигнализации комплекс возбужденных структур мозга, побуждающий к поиску факторов внешней среды, удовлетворяющих потребность, называется мотивацией.Мотивация входит в центральное звено детельности функциональной системы. Именно на основе мотивационного возбуждения происходит формирование поведенческой реакции, направленной на удовлетворение исходной потребности.
В деятельности центров, охваченных мотивационным возбуждением, можно выделить несколько стадий. Начальная и наиболее ответственная- стадия афферентного синтеза.В центрах мозга подвергается оценке не только пусковая, но и обстановочная афферентация. Последняя несет сведения о событиях во внешней среде, и на основе памяти происходит сопоставление внешних раздражителей с внутренними сведениями о них. Факторы внешней среды могут существенно влиять на мотивацию вплоть до полной отмены намечавшегося поведенческого акта. Например, хищник собиравшийся напасть на молодое животное, прекращает эту пищедобывательную реакцию, если в поле его зрения появляется взрослое животное или человек.
Синтез пусковых и обстановочных эфферентных импульсов, а также информации, извлекаемой из памяти (врожденной и индивидуальной). Позволяет формировать цель к действию. Это важнейший этап в деятельности функциональной системы, поэтому нервные структуры, ответственные за это, были выделены П. К. Анохиным как специальный аппарат, названный акцептором результатов действия (АРД; акцептировать означает «принимать решение»). Это аппарат мозга, программирующий результаты будущих событий в виде внутренней психической модели цели (ВМЦ), которая является опережающим отражением реальных событий действительности.
Генетический и индивидуальный опыт подсказывает животному программу действий (ПД): что и в какой последовательности следует делать.
Готовность реализовать действие в оптимальный отрезок времени определяется на основе сопоставления внутреннего субъективного переживания потребности ( величина амплитуды отклонения константы) и появления во внешней среде сигнальных ключевых раздражителей, роль которых глубоко исследована этологами.
Итак, стадия афферентного синтеза заканчивается принятием решения действовать, а воспринятый обстановочный пусковой (ключевой) раздражитель обусловливает выход возбуждения на эфферентные пути. Срабатывает блок выхода системы, т.е. совершаются действия, в результате которых достигается какой то результат, это вторая стадия в деятельности системы. Следующая, третья, стадия ее деятельности заключается в том, что акцептор результатов действия осуществляет сличение параметров результата с качествами (параметрами) внутренней модели цели и внутренней программой действий. Это происходит благодаря потоку обратной импульсации (обратная или вторичная афферентация), возникающей в рецепторах работающих органов, исполняющих поведенческую реакцию. Если сравниваемые параметры совпадают (при этом неизбежно константа возвращается к оптимальному уровню), то поведенческая реакция заканчивается, так как цель достигнута и приспособительный результат получен. В том случае, когда такого совпадения нет, реализуется следующая, четвертая, стадия деятельности функциональной системы. Акцептор результатов действия (АРД) производит перестройку афферентного синтеза и уточнение внутренней модели цели и программы действий. Это становится возможным благодаря возникновению условий, запускающих эмоцию, к которым относится наличие потребности и дефицита информации о дальнейших действиях. Эмоция, возникнув тонизирует мозг, активизирует память и ориентировочно- исследовательскую деятельность животного. Извлекаемые из кладовых памяти дополнительные сведения позволяют уточнить программу действий, обеспечивают в дальнейшем приспособительный эффект и способствуют накоплению индивидуального опыта.
Это движение возбуждения по кольцу от рецепторов работающих органов к центрам, входящим в структуру АРД, может быть многократным и длительным. И все это время поведенческие реакции животного либо недостаточно координированы и требуют усовершенствования, либо достигается некий промежуточный результат, не способствующий возвращению константы к оптимальному уровню.
Обратная афферентация как частный случай обратных связей в биосистемах играет важную роль в активном познавании окружающей среды. На ее основе решается, достигнута ли цель, выполнена ли потребность и какие факторы окружающей среды способствовали этому.
studfiles.net







