Внешние факторы инстинктивного поведения. Какова роль поведенческих реакций животных на факторы внешней среды
Внешние факторы инстинктивного поведения.
Когда говорят об автономности внутренних факторов поведения, об их независимости от внешней среды, то необходимо помнить, что эта независимость является лишь относительной. Уже из приведенных опытов Хольста видно, что эндогенная активность не существует ни «сама по себе», ни «сама для себя»: значение этих спонтанных процессов в центральной нервной системе состоит в прениспозициик возникновению жизненно важных ситуаций («в случае чего – все готово»). В результате животное способно по первому же сигналу незамедлительно и с максимальной пользой для себя реагировать на изменение в окружающей среде.
Эта готовность обеспечивается тем, что соответствующие эндогенные системы периодически активируются как собственной ритмикой, так и внешними воздействиями (например, изменением продолжительности светового дня, повышением или уменьшением температуры и т.д.). Однако инстинктивные движения, по этологической концепции, заблокированы специальной системой «врожденных пусковых механизмов». Последние представляют собой совокупность нейросенсорных систем, обеспечивающих приуроченность поведенческих актов к биологически адекватным условиям среды (к «пусковой ситуации»). Как только животное оказывается в такой ситуации, соответствующий врожденный пусковой механизм обеспечивает распознавание, оценку и интеграцию специфических для данной инстинктивной реакции раздражителей, после чего наступает растормаживание, снятие «блокировки». Очевидно, одновременно происходит активация соответствующих нервных центров и снижение порогов их раздражимости.
Характерной особенностью для врожденных пусковых механизмов является избирательность реагирования на внешние стимулы: они отзываются только на совершенно определенные комбинации раздражителей, которые только и могут вызвать биологически целесообразную реакцию. Иными словами, в сенсорной сфере существует некая «фильтрующая» функция, выражающаяся в специфической преадаптационной «готовности» воспринимать такие раздражители.
Итак, благодаря врожденным пусковым механизмам внутренняя мотивация поведения получает «выход наружу», т.е. создается возможность без индивидуального опыта в биологически значимых ситуациях реагировать так, чтобы это способствовало сохранению особи и вида.
Подводя итог сказанному, можно сказать, что под врожденным пусковым механизмом следует понимать совокупность нейросенсорных систем, обеспечивающих адекватность поведенческих актов по отношению к «пусковой ситуации»: настройку анализаторов на восприятие специфических раздражителей и распознавание последних, интеграцию соответствующих раздражений и растормаживание (или активацию) нервных центров, связанных с данным поведенческим актом.
Внешние раздражители, составляющие в своей совокупности пусковую ситуацию, получили название «ключевых раздражителей», поскольку они подходят к своим врожденным пусковым механизмам, как ключ к замку. Ключевые раздражители являются такими признаками компонентов среды, на которые животные реагируют независимо от индивидуального опыта врожденными, видотипичными формами поведения, точнее, определенными инстинктивными движениями. В описанном поведении курицы это будут определенные общие признаки, свойственные всем ее наземным врагам.
Кроме собственно растормаживающих ключевых раздражителей (их называют также «пусковыми раздражителями») различают также настраивающие ключевые раздражители, предварительно понижающие порог раздражимости нервных центров, причастных к данным действиям животного, а также направляющие ключевые раздражители, о которых пойдет речь при обсуждении таксисов. Общим свойством всех ключевых раздражителей является то, что это специфические элементарные признаки жизненно важных компонентов среды. Ключевыми раздражителями являются простые физические или химические признаки («просто» форма, размер, подвижность, цвет, запах и т.д.), или их пространственные отношения (взаиморасположение деталей, относительная величина и т.д.), или же векторы. Носителями этих признаков могут быть как другие животные, так и растения и объекты неживой природы. В последнем случае ключевые раздражители выполняют преимущественно направляющую функцию. Так, например, немецкий этолог Ф. Вальтер показал, что у детенышей антилоп ключевым стимулом, определяющим выбор места отдыха (лежа, неподвижно), является «что-то вертикальное» вне зависимости от того, что конкретно это за объект.
Наилучшим образом изучены ключевые раздражители, носителями которых являются животные. Эти раздражители представляют особый интерес и потому, что являются первичными, генетически фиксированными элементами общения у животных. Так, в ставших уже классическими опытах голландского зоолога Н. Тинбергена, одного из основоположников современной этологии, с помощью макетов изучалась пищевая реакция («попрошайничество») птенцов серебристых чаек (клевание клюва родительской особи) и дроздов (вытягивание шеи и раскрытие клюва) при появлении родительской особи.
В естественных условиях голодный птенец серебристой чайки клюет красное пятно на желтом клюве родителя, и тот в ответ отрыгивает пищу в рот птенцу. В опытах предъявлялась серия все более упрощаемых моделей – макетов. Первая модель точно воспроизводила внешний облик естественного носителя ключевых раздражителей, т.е. головы взрослой серебристой чайки с желтым клювом и красным пятном на нем. В последующих моделях путем проб постепенно исключались отдельные детали, и в результате макет становился все менее похожим на голову птицы (рис. 7). В конце концов остался лишь плоский красный предмет с продолговатым выступом. Но этот предмет оказался способным вызвать даже более сильную реакцию птенцов, чем исходная модель. Еще более эта реакция может быть усилена, если этот макет заменить тонкой белой палочкой, исчерченной поперечными темно-красными полосами. Ключевыми раздражителями в данном примере будут просто «красное» и «продолговатое».
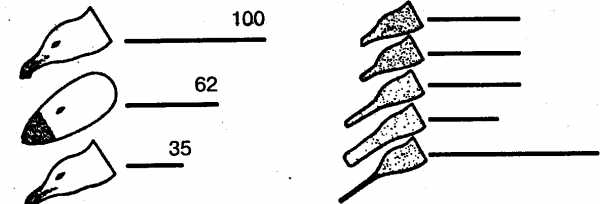
Рис. 7. Опыты Тинбергена с применением макетов головы серебристой чайки. Линиями обозначена интенсивность реакций птенцов на макет (число клеваний).
В опытах, проведенных Тинбергеном с десятидневными птенцами дроздов, выявилась другая категория ключевых раздражителей – взаиморасположение и относительная величина деталей объекта-носителя. В качестве макетов использовались плоские диски. Если показать птенцам такой круг, они будут тянуться к его верхней части (местонахождению головы птиц). Если же к большому кругу прибавить маленький, птенцы потянутся к нему. Если, наконец, прибавить к большому кругу два меньших круга, но разных размеров, то решающее значение приобретет относительная величина этих фигур. Размеры «головы» не должны приближаться к размерам «туловища»: при большом «туловище» птенцы потянутся к большему дополнительному кругу («голове») (рис. 8), при маленьком же–к меньшему.
Интересные опыты проводил еще в середине 30-х годов Г.Л. Скребицкий совместно с Т.И. Бибиковой на подмосковном озере Киёво, изучая отношение чайки к ее яйцам. Яйца перекладывались из гнезда в гнездо, заменялись яйцами других видов, искусственными яйцами, а затем и различными посторонними предметами различной величины, формы и окраски. Оказалось, что чайки садились как на чужие, так и на деревянные, стеклянные, каменные, глиняные яйца различной величины и самой разнообразной окраски и начинали их «высиживать». То же самое наблюдалось, когда вместо яиц в гнездо подкладывались разноцветные шары, камешки или картофелины. По свидетельству экспериментаторов, «чайки, сидящие на таких предметах, представляли очень оригинальную картину, но в особенности необычайным становилось зрелище, когда согнанная с гнезда птица возвращалась к нему обратно и, прежде чем сесть, заботливо поправляла клювом разноцветные шары, камешки или картошку».35
Если, однако, чайкам подкладывались предметы иной формы, например кубики или камни с неровными краями, поведение птиц заметно изменялось. Когда за край гнезда клалось по одному яйцу и инородному предмету, птицы вкатывали обратно в гнездо наряду с яйцами лишь округлые гладкие предметы, размеры которых соответствовали размерам яиц. В известных пределах не препятствовали этому существенные отклонения от нормы в весе (каменное яйцо весило в два с лишним раза больше, чем чаечье), материал, из которого был изготовлен предмет, и окраска.

Исследователи пришли к выводу, что положительная реакция чайки на яйцо определяется лишь несколькими его элементарными признаками: округлостью, отсутствием выступов, углублений или насечек (рис. 9). Именно эти признаки и выступали здесь как ключевые раздражители.
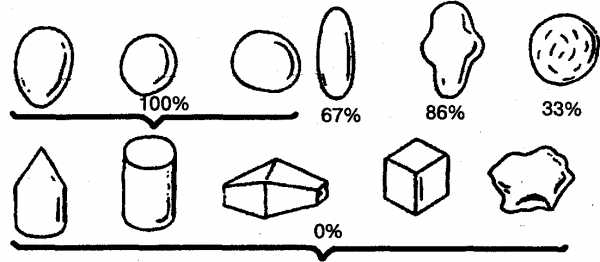
Рис. 9. Вкатывание в гнездо различных предметов в зависимости от их свойств у обыкновенной чайки (в процентах). Верхний ряд, слева направо: яйца (чаечьи, куриные, деревянные и каменные), деревянные шары, другие округлые предметы (картошка, камни) удлиненное глиняное яйцо с углублениями, глиняный шар с зарубками. Нижний ряд, слева направо: деревянный конус, деревянный цилиндр, деревянные и глиняные многоугольники, деревянные и глиняные кубики, угловатые камни (по СкребицкомуиБибиковой).
Важная особенность действия ключевых раздражителей заключается в том, что они подчиняются закону суммации: с увеличением их параметров пропорционально усиливается инстинктивная реакция животного. В экспериментальных условиях это может привести к так называемым «супероптимальным» реакциям, когда животное «преувеличенно», сильнее, чем в норме, реагирует на искусственный раздражитель, в котором «сгущены краски». Мы уже встречались с этим явлением при описании опыта с птенцами чайки, когда они сильнее реагировали на поперечно исчерченную красными полосками палочку, чем на настоящий клюв живой птицы. Суммация проистекает здесь из многократности красных меток и их большей контрастности.
Эффект супероптимальной реакции может в условиях эксперимента привести даже к биологически абсурдному поведению животного. Если, например, предложить чайке два яйца разной величины, она закатит в гнездо более крупное. В итоге может возникнуть такое положение, что птица бросит свое яйцо, чтобы попытаться высиживать деревянный макет яйца гигантских размеров, обладающих супероптимальными признаками ключевого раздражителя.
Как видно из приведенных примеров, ключевые стимулы действуют на поведение животного принудительно, заставляя его выполнять определенные инстинктивные движения, невзирая на возможно воспринимаемую животным общую ситуацию. Это объясняет многие, ранее казавшиеся загадочными моменты в поведении животных.
Так, например, еще в начале нашего века один из авторов антропоморфических сочинений по психологии животных Т. Целл дал следующий ответ на вопрос о том, почему крупные хищники в нормальных условиях при встрече с человеком не нападают на него: например, лев не нападает не потому, что почитает человека, а потому, что не уверен в исходе схватки. «Есть ли у человека оружие, да какое оно, это оружие? – думает лев. – Да ну его, пойду-ка я лучше своей дорогой».
Конечно, лев, как и другие крупные хищники, как правило, уклоняется от нападения на человека не потому, что руководствуется такими глубокомысленными рассуждениями. Разгадка «почтительного» отношения диких зверей к человеку, очевидно, кроется в следующем. Будучи сытым, хищник не реагирует и на присутствие животных, которыми обычно питается. У голодного же хищника преследование животных-жертв и нападение на них обусловливаются сочетанием рассмотренных выше внутренних факторов (первопричина – изменение уравновешенности внутренней среды организма в результате недостатка питательных веществ) с соответствующими внешними стимулами – ключевыми раздражителями, носителями которых являются естественные объекты питания, т.е. животные-жертвы, но не человек.
Даже самый кровожадный хищник не волен нападать на кого угодно и когда угодно. Эти действия также ориентируются во времени и пространстве ключевыми раздражителями, как и все прочие поведенческие акты. Другими словами, дело не в том, «хочет» или «не хочет» животное поступить так или иначе. Если внутреннее состояние животного соответствует определенной внешней пусковой ситуации, то оно волей-неволей вынуждено вести себя так, как это диктует для данных условий генетически зафиксированный код видотипичного поведения.
* * *
Итак, в процессе эволюции возникают адаптации к более постоянным компонентам внешней среды, необходимым для удовлетворения потребностей, непрестанно возникающих в результате изменений внутренней среды организма. Нахождение (или избегание) важных для организма компонентов внешней среды осуществляется путем ориентации по типичным признакам этих компонентов – ключевым раздражителям.
Результаты этой ориентации реализуются нейросенсорными системами (врожденными пусковыми механизмами), которые действуют рефлекторно и включают эндогенные, генетически фиксированные компоненты инстинктивного поведения. Таким образом, действуя «вовне», врожденные пусковые механизмы обеспечивают избирательную направленность внешней активности организма лишь на определенные сигнальные стимулы; действуя же «вовнутрь», они осуществляют оценку и отбор поступающей через рецепторы информации и ее реализацию для активации или понижения порогов раздражимости соответствующих нервных структур, для снятия «блокировки», растормаживания эндогенных нервных процессов, мотивирующих инстинктивные движения и действия. Таким образом осуществляется на врожденной основе корреляция внутренних потребностей организма с биологически существенными изменениями в окружающей его среде.
Только в этой корреляции состоит все биологическое значение эндогенной мотивации поведения. Внутренние стимулы служат лишь для осуществления движения по отношению к среде, без которого организм – именно как саморегулирующаяся система – нежизнеспособен. И в этом, широком, смысле деятельность всей нервной системы в целом всегда рефлекторна.
Равным образом даже наиболее как будто «далеко упрятанные» от внешней среды факторы поведения сами зависят от процессов обмена веществ. Эти процессы уже непосредственно связаны с окружающей организм средой. А так как организм активно регулирует, создает необходимые внешние предпосылки для нормального протекания обменных процессов как раз посредством поведения, то круг смыкается. И в этом отношении проявляется относительность независимости эндогенных автоматизмов, спонтанной активности нервной системы, способности к «самопрограммированию». Ясно, что все эти эндогенные процессы лишь постольку независимы от внешней среды, поскольку они лишь опосредованно связаны с ней.
studfiles.net
Внешние факторы инстинктивного поведения
Когда говорят об автономности внутренних факторов поведения, об их независимости от внешней среды, то необходимо помнить, что эта независимость является лишь относительной. Уже из приведенных опытов Хольста видно, что эндогенная активность не существует ни «сама по себе», ни «сама для себя»: значение этих спонтанных процессов в центральной нервной системе состоит в прениспозиции к возникновению жизненно важных ситуаций («в случае чего – все готово»). В результате животное способно по первому же сигналу незамедлительно и с максимальной пользой для себя реагировать на изменение в окружающей среде.
Эта готовность обеспечивается тем, что соответствующие эндогенные системы периодически активируются как собственной ритмикой, так и внешними воздействиями (например, изменением продолжительности светового дня, повышением или уменьшением температуры и т.д.). Однако инстинктивные движения, по этологической концепции, заблокированы специальной системой «врожденных пусковых механизмов». Последние представляют собой совокупность нейросенсорных систем, обеспечивающих приуроченность поведенческих актов к биологически адекватным условиям среды (к «пусковой ситуации»). Как только животное оказывается в такой ситуации, соответствующий врожденный пусковой механизм обеспечивает распознавание, оценку и интеграцию специфических для данной инстинктивной реакции раздражителей, после чего наступает растормаживание, снятие «блокировки». Очевидно, одновременно происходит активация соответствующих нервных центров и снижение порогов их раздражимости.
Характерной особенностью для врожденных пусковых механизмов является избирательность реагирования на внешние стимулы: они отзываются только на совершенно определенные комбинации раздражителей, которые только и могут вызвать биологически целесообразную реакцию. Иными словами, в сенсорной сфере существует некая «фильтрующая» функция, выражающаяся в специфической преадаптационной «готовности» воспринимать такие раздражители.
Итак, благодаря врожденным пусковым механизмам внутренняя мотивация поведения получает «выход наружу», т.е. создается возможность без индивидуального опыта в биологически значимых ситуациях реагировать так, чтобы это способствовало сохранению особи и вида.
Подводя итог сказанному, можно сказать, что под врожденным пусковым механизмом следует понимать совокупность нейросенсорных систем, обеспечивающих адекватность поведенческих актов по отношению к «пусковой ситуации»: настройку анализаторов на восприятие специфических раздражителей и распознавание последних, интеграцию соответствующих раздражений и растормаживание (или активацию) нервных центров, связанных с данным поведенческим актом.
Внешние раздражители, составляющие в своей совокупности пусковую ситуацию, получили название «ключевых раздражителей», поскольку они подходят к своим врожденным пусковым механизмам, как ключ к замку. Ключевые раздражители являются такими признаками компонентов среды, на которые животные реагируют независимо от индивидуального опыта врожденными, видотипичными формами поведения, точнее, определенными инстинктивными движениями. В описанном поведении курицы это будут определенные общие признаки, свойственные всем ее наземным врагам.
Кроме собственно растормаживающих ключевых раздражителей (их называют также «пусковыми раздражителями») различают также настраивающие ключевые раздражители, предварительно понижающие порог раздражимости нервных центров, причастных к данным действиям животного, а также направляющие ключевые раздражители, о которых пойдет речь при обсуждении таксисов. Общим свойством всех ключевых раздражителей является то, что это специфические элементарные признаки жизненно важных компонентов среды. Ключевыми раздражителями являются простые физические или химические признаки («просто» форма, размер, подвижность, цвет, запах и т.д.), или их пространственные отношения (взаиморасположение деталей, относительная величина и т.д.), или же векторы. Носителями этих признаков могут быть как другие животные, так и растения и объекты неживой природы. В последнем случае ключевые раздражители выполняют преимущественно направляющую функцию. Так, например, немецкий этолог Ф. Вальтер показал, что у детенышей антилоп ключевым стимулом, определяющим выбор места отдыха (лежа, неподвижно), является «что-то вертикальное» вне зависимости от того, что конкретно это за объект.
Наилучшим образом изучены ключевые раздражители, носителями которых являются животные. Эти раздражители представляют особый интерес и потому, что являются первичными, генетически фиксированными элементами общения у животных. Так, в ставших уже классическими опытах голландского зоолога Н. Тинбергена, одного из основоположников современной этологии, с помощью макетов изучалась пищевая реакция («попрошайничество») птенцов серебристых чаек (клевание клюва родительской особи) и дроздов (вытягивание шеи и раскрытие клюва) при появлении родительской особи.
В естественных условиях голодный птенец серебристой чайки клюет красное пятно на желтом клюве родителя, и тот в ответ отрыгивает пищу в рот птенцу. В опытах предъявлялась серия все более упрощаемых моделей – макетов. Первая модель точно воспроизводила внешний облик естественного носителя ключевых раздражителей, т.е. головы взрослой серебристой чайки с желтым клювом и красным пятном на нем. В последующих моделях путем проб постепенно исключались отдельные детали, и в результате макет становился все менее похожим на голову птицы (рис. 7). В конце концов остался лишь плоский красный предмет с продолговатым выступом. Но этот предмет оказался способным вызвать даже более сильную реакцию птенцов, чем исходная модель. Еще более эта реакция может быть усилена, если этот макет заменить тонкой белой палочкой, исчерченной поперечными темно-красными полосами. Ключевыми раздражителями в данном примере будут просто «красное» и «продолговатое».
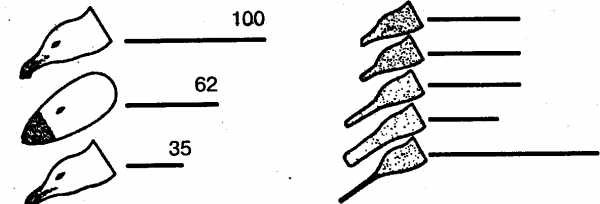
Рис. 7. Опыты Тинбергена с применением макетов головы серебристой чайки. Линиями обозначена интенсивность реакций птенцов на макет (число клеваний)
В опытах, проведенных Тинбергеном с десятидневными птенцами дроздов, выявилась другая категория ключевых раздражителей – взаиморасположение и относительная величина деталей объекта-носителя. В качестве макетов использовались плоские диски. Если показать птенцам такой круг, они будут тянуться к его верхней части (местонахождению головы птиц). Если же к большому кругу прибавить маленький, птенцы потянутся к нему. Если, наконец, прибавить к большому кругу два меньших круга, но разных размеров, то решающее значение приобретет относительная величина этих фигур. Размеры «головы» не должны приближаться к размерам «туловища»: при большом «туловище» птенцы потянутся к большему дополнительному кругу («голове») (рис. 8), при маленьком же–к меньшему.
Интересные опыты проводил еще в середине 30-х годов Г.Л. Скребицкий совместно с Т.И. Бибиковой на подмосковном озере Киёво, изучая отношение чайки к ее яйцам. Яйца перекладывались из гнезда в гнездо, заменялись яйцами других видов, искусственными яйцами, а затем и различными посторонними предметами различной величины, формы и окраски. Оказалось, что чайки садились как на чужие, так и на деревянные, стеклянные, каменные, глиняные яйца различной величины и самой разнообразной окраски и начинали их «высиживать». То же самое наблюдалось, когда вместо яиц в гнездо подкладывались разноцветные шары, камешки или картофелины. По свидетельству экспериментаторов, «чайки, сидящие на таких предметах, представляли очень оригинальную картину, но в особенности необычайным становилось зрелище, когда согнанная с гнезда птица возвращалась к нему обратно и, прежде чем сесть, заботливо поправляла клювом разноцветные шары, камешки или картошку».*
Скребицкий Г. Отношение чайки к гнезду, яйцам и птенцам // Рефлексы, инстинкты и навыки. М., 1936. С. 144–145.
Если, однако, чайкам подкладывались предметы иной формы, например кубики или камни с неровными краями, поведение птиц заметно изменялось. Когда за край гнезда клалось по одному яйцу и инородному предмету, птицы вкатывали обратно в гнездо наряду с яйцами лишь округлые гладкие предметы, размеры которых соответствовали размерам яиц. В известных пределах не препятствовали этому существенные отклонения от нормы в весе (каменное яйцо весило в два с лишним раза больше, чем чаечье), материал, из которого был изготовлен предмет, и окраска.

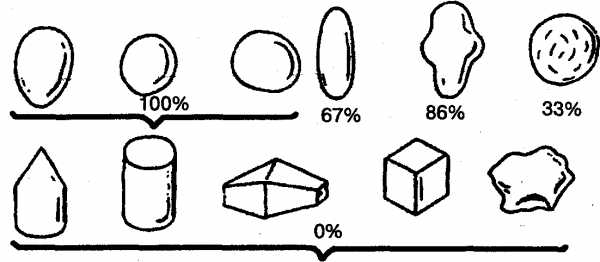
Рис. 9. Вкатывание в гнездо различных предметов в зависимости от их свойств у обыкновенной чайки (в процентах). Верхний ряд, слева направо: яйца (чаечьи, куриные, деревянные и каменные), деревянные шары, другие округлые предметы (картошка, камни) удлиненное глиняное яйцо с углублениями, глиняный шар с зарубками. Нижний ряд, слева направо: деревянный конус, деревянный цилиндр, деревянные и глиняные многоугольники, деревянные и глиняные кубики, угловатые камни (по Скребицкому и Бибиковой)
Важная особенность действия ключевых раздражителей заключается в том, что они подчиняются закону суммации: с увеличением их параметров пропорционально усиливается инстинктивная реакция животного. В экспериментальных условиях это может привести к так называемым «супероптимальным» реакциям, когда животное «преувеличенно», сильнее, чем в норме, реагирует на искусственный раздражитель, в котором «сгущены краски». Мы уже встречались с этим явлением при описании опыта с птенцами чайки, когда они сильнее реагировали на поперечно исчерченную красными полосками палочку, чем на настоящий клюв живой птицы. Суммация проистекает здесь из многократности красных меток и их большей контрастности.
Эффект супероптимальной реакции может в условиях эксперимента привести даже к биологически абсурдному поведению животного. Если, например, предложить чайке два яйца разной величины, она закатит в гнездо более крупное. В итоге может возникнуть такое положение, что птица бросит свое яйцо, чтобы попытаться высиживать деревянный макет яйца гигантских размеров, обладающих супероптимальными признаками ключевого раздражителя.
Как видно из приведенных примеров, ключевые стимулы действуют на поведение животного принудительно, заставляя его выполнять определенные инстинктивные движения, невзирая на возможно воспринимаемую животным общую ситуацию. Это объясняет многие, ранее казавшиеся загадочными моменты в поведении животных.
Так, например, еще в начале нашего века один из авторов антропоморфических сочинений по психологии животных Т. Целл дал следующий ответ на вопрос о том, почему крупные хищники в нормальных условиях при встрече с человеком не нападают на него: например, лев не нападает не потому, что почитает человека, а потому, что не уверен в исходе схватки. «Есть ли у человека оружие, да какое оно, это оружие? – думает лев. – Да ну его, пойду-ка я лучше своей дорогой».
Конечно, лев, как и другие крупные хищники, как правило, уклоняется от нападения на человека не потому, что руководствуется такими глубокомысленными рассуждениями. Разгадка «почтительного» отношения диких зверей к человеку, очевидно, кроется в следующем. Будучи сытым, хищник не реагирует и на присутствие животных, которыми обычно питается. У голодного же хищника преследование животных-жертв и нападение на них обусловливаются сочетанием рассмотренных выше внутренних факторов (первопричина – изменение уравновешенности внутренней среды организма в результате недостатка питательных веществ) с соответствующими внешними стимулами – ключевыми раздражителями, носителями которых являются естественные объекты питания, т.е. животные-жертвы, но не человек.
Даже самый кровожадный хищник не волен нападать на кого угодно и когда угодно. Эти действия также ориентируются во времени и пространстве ключевыми раздражителями, как и все прочие поведенческие акты. Другими словами, дело не в том, «хочет» или «не хочет» животное поступить так или иначе. Если внутреннее состояние животного соответствует определенной внешней пусковой ситуации, то оно волей-неволей вынуждено вести себя так, как это диктует для данных условий генетически зафиксированный код видотипичного поведения.
* * *
Итак, в процессе эволюции возникают адаптации к более постоянным компонентам внешней среды, необходимым для удовлетворения потребностей, непрестанно возникающих в результате изменений внутренней среды организма. Нахождение (или избегание) важных для организма компонентов внешней среды осуществляется путем ориентации по типичным признакам этих компонентов – ключевым раздражителям.
Результаты этой ориентации реализуются нейросенсорными системами (врожденными пусковыми механизмами), которые действуют рефлекторно и включают эндогенные, генетически фиксированные компоненты инстинктивного поведения. Таким образом, действуя «вовне», врожденные пусковые механизмы обеспечивают избирательную направленность внешней активности организма лишь на определенные сигнальные стимулы; действуя же «вовнутрь», они осуществляют оценку и отбор поступающей через рецепторы информации и ее реализацию для активации или понижения порогов раздражимости соответствующих нервных структур, для снятия «блокировки», растормаживания эндогенных нервных процессов, мотивирующих инстинктивные движения и действия. Таким образом осуществляется на врожденной основе корреляция внутренних потребностей организма с биологически существенными изменениями в окружающей его среде.
Только в этой корреляции состоит все биологическое значение эндогенной мотивации поведения. Внутренние стимулы служат лишь для осуществления движения по отношению к среде, без которого организм – именно как саморегулирующаяся система – нежизнеспособен. И в этом, широком, смысле деятельность всей нервной системы в целом всегда рефлекторна.
Равным образом даже наиболее как будто «далеко упрятанные» от внешней среды факторы поведения сами зависят от процессов обмена веществ. Эти процессы уже непосредственно связаны с окружающей организм средой. А так как организм активно регулирует, создает необходимые внешние предпосылки для нормального протекания обменных процессов как раз посредством поведения, то круг смыкается. И в этом отношении проявляется относительность независимости эндогенных автоматизмов, спонтанной активности нервной системы, способности к «самопрограммированию». Ясно, что все эти эндогенные процессы лишь постольку независимы от внешней среды, поскольку они лишь опосредованно связаны с ней.
studfiles.net
Этология
Применение учения И. П. Павлова и этологии в животноводстве
ЭТОЛОГИЯ — НАУКА
О ПОВЕДЕНИИ ЖИВОТНЫХ
Термин «этология» происходит и греческого слова «этос» и означает поведение, характер. Поведение- это совокупность проявлений внешней, преимущественно двигательной активности животного, необходимых для связи организма со средой. Этология как наука о биологических закономерностях поведения значительное развитие получила лишь на рубеже XIX и XX столетий, но наблюдение за поведением животных велось с давних пор.
Одним из родоначальников этологии считают Ч. Дарвина (1809—1882), опубликовавшего труд «Выражение эмоций у животных и человека» (1882), в котором он утверждал, что человек и животные обладают весьма сходными чувствами, инстинктами и эмоциями.
Значительным этапом в науке о поведении было появление в конце прошлого века нового направления — бихевиоризма (от английского слова «behavior» —поведение). Основоположник бихевиоризма американский психолог Э. Торндайк (1874—1949) изучал поведение цыплят, кошек, собак, обезьян объективным методом. Животное помещали в ящик, и оно могло выйти из него к пище или на свободу, выучившись открывать дверцу. Торндайк обратил внимание на связь между стимулом и реакцией как основу поведения животных. Его последователи усложнили эти эксперименты, использовав лабиринтные методики. Был накоплен большой и интересный материал относительно скорости обучения различных животных, длительности сохранения навыков и т. д. Однако бихевиори-сты, проводя эксперименты, не обращали внимания на самое главное — на мозговые процессы, возникающие в результате действия стимула, вследствие которых и развивается ответная деятельность организма.
Иной подход к изучению психических явлений связан с направлением, получившим название гештальтпсихологии. Один из его основоположников Р. Келлер (1887—1967) изучал поведение шимпанзе в условиях, в которых они могли научиться применять «орудия» (палки и т. д.), чтобы достать пищу, находившуюся в клетке или подвешенную к потолку. Анализируя опыты, он пришел к выводу, что шимпанзе обладают разумной деятельностью типа человеческой. С точки зрения гештальтистов, психике изначально присуще свойство образовывать образы (гештальты). Но хотя они и критиковали бихевиористов за механицизм, сами также не пытались связать свои представления с конкретными механизмами мозговой деятельности, как это сделал И. П. Павлов.
И. П. Павлов и его последователи изучали физиологические механизмы, лежащие в основе высшей нервной деятельности,— условные рефлексы. Но вместе с тем они признавали огромную важность полного изучения всех нервных реакций организма, лежа- щих в основе адаптивного поведения животных. Наши знания о врожденных, генетических, безусловных рефлексах (инстинктах) очень малы. Павлов считал, что инстинкты лежат в основе формирования индивидуального приобретенного поведения. Но любой акт поведения является и реакцией на внешние раздражения. Следовательно, все поведенческие реакции формируются при участии генетических факторов и под влиянием внешней среды.
Начиная с конца XIX в. ученые стали исследовать общее поведение животных, как врожденное, так и приобретенное. Первая работа была проведена и опубликована в 1894 г. Л. Морганом, наблюдавшим за поведением своей собаки. Обобщенные результаты своих исследований он опубликовал в книге «Привычка и инстинкт», вышедшей в русском переводе в 1899 г.
В 20—30-х годах XX в. сложилась так называемая объективистская школа, которая основное внимание уделяла наблюдению в естественных условиях, вне стен лаборатории. Выдающиеся ее представители — К. Лоренц, Н. Тинберген, К. Фриш изучали инстинктивное поведение животных и его развитие в онто- и филогенезе. Их считают основоположниками этологии.
Существенный вклад в науку о поведении животных вносят советские ученые. Элементарной рассудочной деятельности животных посвящены работы Л. В. Крушинского, поведению птиц — А. Н. Промптова; особенности поведения млекопитающих, обусловленных запаховыми сигналами (феромонами), изучаются под руководством В. Е. Соколова; широко известны работы А. Д. Слонима, исследовавшего поведенческие реакции животных в различных условиях существования; Л. М. Баскин изучает вопросы поведения копытных; проводится много других исследований.
В этологии поведение рассматривается как многообразное взаимодействие животного с окружающей средой. В результате наблюдения за организмом устанавливается в основном внешнее проявление его реакций. Физиология изучает нервные механизмы деятельности мозга, обеспечивающие поведение животных. Этологами, в основном зоологами, собраны многочисленные материалы, характеризующие поведение многих видов животного мира.
Применение в практике животноводства результатов научных исследований поведения сельскохозяйственных животных позволяет значительно повысить их продуктивность
в результате более целесообразного, экономичного содержания, кормления и разведения.
Формы поведения. Поведение включает в себя любой вид активности, проявляемой индивидуумом. Для характеристики .поведения Л. В. Крушинский (1960) предложил два понятия: «унитарная реакция» и «биологическая форма поведения». Унитарная реакция — это целостный акт поведения, формирующийся в результате объединения (интеграции) условных и безусловных рефлексов, соотношение которых может иметь некоторые вариации. Данный акт направлен на выполнение одиночного приспособительного действия, которое при различных способах своего осуществления имеет определенный шаблон. Унитарная реакция — это элементарная частица поведения. На основе интеграции указанных поведений образуются «биологические формы поведения» животных. Эти многоактные поведения обеспечивают основные биологические потребности организма. Например, для собаки характерно лакать воду и жидкий корм стоя, с опущенным хвостом. Твердую пищу она кусает и жует леж<а, придерживая кусок передними лапами и отрывая зубами от него небольшие части.
Виды поведений классифицируют по приспособительному эффекту. Различают поведения: половое, родительское, пищевое, оборонительное, исследовательское, доминирование и т. д.
На различных этапах эволюции можно выделить следующие врожденные адаптивные реакции: таксисы, рефлексы и инстинкты. К приобретенным формам, более изменчивым, относят обучение и мышление.
Таксисы — простейшая форма поведения, определяющая взаимодействие организма со средой у простейших и многоклеточных. На последующих ступенях эволюции роль таксисов резко падает и они заменяются другими, более совершенными механизмами адаптации. Таксисы представляют собой ориентацию по отношению к некоторым факторам среды. В простейшем случае таксис является ориентацией или движением', в котором адаптация организма есть простой врожденный автоматический ответ на стимул. В других случаях таксис может быть лишь элементом сложного поведения.
Рефлекс — тоже вид адаптивного поведения. Здесь он рассматривается как безусловнорефлекторная реакция, служащая одним из главных видов адаптации в животном мире
Инстинкт — более высшая форма врожденного поведения, сформировавшаяся на протяжении истории вида. Это наследственные комплексы реакций на определенные воздействия. Инстинктивное поведение, как и все другие формы поведения, имеет определенную направленность — всегда служить целям сохранения и развития организма 5 условиях, характерных для жизни этого вида животных. У высших животных трудно выделить в поведении врожденные элементы и быть уверенным, что обучение не повлиялс на поведение. Например, у кошег ловля мышей считается инстинктивной реакцией, но часто котята прежде чем научиться ловить мышей должны увидеть, как это делан: взрослые кошки. Чисто безусловно-рефлекторным поведенческий ак~ может быть только в первый р.; в жизни, а затем на него наслаивается масса условных рефлексе
Обучение — процесс, благодаря которому жизненный опыт влияет г-поведение каждого индивидуума -который позволяет животному ра:-вить новые приспособительные peaкции с учетом прошлого опыта, i также видоизменять те реакции, которые оказались неадаптивными.;
Существует много видов обучения, варьирующих от простейших модификаций врожденного поведения до сложнейших процессов, свойственных умственной деятельности человека. Приводим некоторые из них.
Запечатление - импринтинг. Понятие о запечатлении впервые было дано К. Лоренцом (1937). Это явление впервые было описано у птиц, впоследствии его обнаружили у овец, коз, оленей, лошадей и других животных, детеныши которых сразу после рождения способны передвигаться (зрелорождающиеся). У птенцов или детенышей возникает особая «привязанность» к первому движущемуся объекту, который они увидели, и они реагируют на него так, как если бы это была их мать.
Запечатление заключается в установлении характерной связи животного с объектом внешней среды в определенный период его жизни. Эта связь может проявляться в следовании за любым движущимся объектом, в приближении к этому объекту, вступлении с ним в контакт, прикосновении, лизании, подаче звуков низкого тона (звуков довольства), высокого тона (дистресс-реакции). Примером этого может служить поведение только что родившегося ягненка. Если в данный момент удалить его от матери и стать перед ним, а затем пойти, ягненок будет двигаться за человеком.
Объектом запечатления на протяжении последующей жизни могут быть и другие предметы и явления, как-то: особо опасные предметы, тревожные звуки и пр. Импринтинг в корне отличается от условного рефлекса, так как очень быстро запоминается и реакция на запечатленный объект длительно сохраняется, образуется он в определенный «критический» период развития жизни и не угасает (К. Лоренц, 1971). Однако эти отличия не абсолютны. Импринтинг имеет- большое значение в формировании поведения животного, при этом быстро и прочно запоминаются важные объекты и
явления. В истолковании механизма явления импринтинга нет единства мнений. Некоторые исследователи причисляют импринтинг к одной из форм ранней памяти (А. Г. Понугаева, 1973), другие относят импринтинг к суммационным рефлексам — самой простой форме обучения, основанной на процессах сенсибилизации (Л. Г. Воронин, 1969).
Одна из наиболее изученных форм поведения — условный рефлекс, открытый и разработанный И. П. Павловым. Метод условных рефлексов при изучении поведения животных самый плодотворный и объективный.
Условная реакция — это сложный адаптивный ответ животного на определенный условный раздражитель. Она возникает в том случае, если вслед за условным раздражителем дается безусловный. Условный рефлекс— основная форма обучения*
Некоторыми зарубежными исследователями школы бихевиористов рефлекторная теория И. П. Павлова трактуется по схеме «стимул — реакция», что, по существу, может быть распространено весьма ограниченно на рефлекторную деятельность низшего уровня. Схема «стимул — реакция» не отображает всей сложности структуры даже безусловного рефлекса, не говоря уже о дугах условного рефлекса, где связь между раздражителем и реакцией непостоянна.
Отличие инструментального условного рефлекса от так называемого классического условного рефлекса состоит в том, что индифферентный раздражитель, например звонок, подкрепляется пищей не каждый раз, а только в том случае, если животное нажимает на рычаг. Но если однажды эта реакция будет выполнена при включении звонка, а за ней сразу последует подкрепление пищей, то вероятность того, что животное вновь нажмет на рычаг, увеличится. Примером инструментального условного рефлекса может служить процесс питья воды из автопоилки. Животное нажимает мордой клапан, вода поступает в резервуар поилки, и корова пьет. В данном рефлексе выявляются причинно-следственные отношения, а факт безусловного подкрепления зависит от действия самого животного.
Метод проб и ошибок — усложненный инструментальный рефлекс, при котором задача решается в результате слепого поиска.
Подражание — одна из форм обучения. Случаи подражания у животных наблюдают очень часто. Иногда подражание происходит в виде автоматических реакций, близких к импринтингу — рефлексу следования. При погрузке лошадей в вагоны некоторые из них сопротивляются и не входят в вагон. Но стоит лишь на глазах такой лошади ввести другую, спокойно входящую в вагон, как вслед за ней строптивая лошадь входит значительно спокойнее. Кроме освоения полезных рефлексов, животные путем подражания могут выработать у себя различные дурные привычки.
К видам обучения относится инсайт — внезапное осуществление новой реакции без предварительных проб и ошибок. Пример инсайта - использование животными орудий.
Так, шимпанзе ставят ящики один на другой или соединяют две палки, чтобы достать высоко подвешенный банан, и этими приемами пользуются без предварительного обучения. Инсайт можно рассматривать как выражение способности к образному мышлению.
Мышление — высшая форма поведения, доминирующая у человека. У высших животных доказано наличие элементарной рассудочной деятельности. Примером может служить инсайт. Иногда после ряда неудачных попыток и наступившей затем паузы животное внезапно изменяет тактику своего поведения и решает задачу. Следовательно, в мозге животного произошла оценка ранее предпринятых попыток и внесен корректив в план дальнейших действий.
У высших животных существуют и в эволюционном плане развиваются элементы рассудочной деятельности. Это доказывается решением животными сложных задач.
Рассмотренные формы приобретенного поведения — обучение и мышление — возникают на высших ступенях эволюции. Обучение становится доминирующим у млекопитающих. Поведение их определяется реакциями, врожденными и приобретенными в результате обучения.
Формирование поведения животных.
У лошадей, крупного и мелкого рогатого скота и свиней молодняк рождается настолько зрелым, что может следовать за кормящей самкой иногда на довольно большие расстояния. Развитие его в естественных условиях происходит в табуне и стаде, совершающих ежедневно значительные переходы по пастбищам.
Эмбриональное развитие у этих животных протекает таким образом, что новорожденному обеспечена возможность выживания при соответствующих условиях существования. У копытных забота о выживании новорожденных в значительной мере предоставлена самим животным. Однако и то обстоятельство, что новорожденный может самостоятельно ходить, не устраняет необходимости наблюдения за ним, создания условий, адекватных для его нормального развития.
Для организации правильного развития молодняка необходимы знание всех потребностей новорожденных, учет особенностей формирования их поведения, обучения и приобретения ими навыков, необходимых для жизни. Поведение взрослого животного в каждый данный момент, его реакции на те или иные раздражители в значительной мере зависят от истории формирования его поведения в молодом возрасте.
У самок еще до родов выделяемый гипофизом гормон пролактин вызывает материнскую доминанту — проявление инстинкта материнства, обеспечивающего выращивание и защиту приплода. Самки, находящиеся вне помещения (в табуне, отаре, стаде), перед родами удаляются от стада, выбирают укромное место, где и происходят роды. Если не подготовлено логово или кругом очень шумно, роды могут задержаться. Они обычно проходят в ночное время, когда в помещении тихо и спокойно. Тотчас после родов самки беспокоятся, часто кричат, обнюхивают новорожденных и начинают их облизывать. Облизывание, продолжающееся длительное время, иногда часами, массирует кожу, что способствует нормальному кровообращению. Считают, что в первые часы после родов самки запоминают запах и облик своих детенышей. С первых же часов после родов самки становятся агрессивными по отношению к другим животным и людям. Главная роль в охране детенышей принадлежит матери, причем при защите потомства родители часто проявляют самоотверженность.
В поведении новорожденных уже в первые часы или даже минуты после рождения можно проследить ряд врожденных рефлексов. Пищевой рефлекс у млекопитающих проявляется в виде сосания. Новорожденные тянутся к соскам матери и активно высасывают молозиво. При этом они энергично подталкивают вымя, способствуя выделению молока.
Стимулом появления сосательного рефлекса у телят, жеребят и особенно у ягнят служит затемнение, когда мать станет над детенышем. Это затемнение вызывает реакцию ягненка, он поднимает морду кверху, касается шерсти брюха и находит соски.
Уже через несколько минут после рождения новорожденные начинают проявлять настороженность, пугливость. И. П. Павлов назвал этот страх первичным, временным рефлексом биологической осторожности. Пугливость и осторожность обычно дольше сохраняются у новорожденных, содержащихся изолированно, чем у содержащихся вместе и усваивающих опыт других животных. Страх может возникнуть и в последующие этапы развития животного при изменении обстановки, появлении новой одежды у пастухов, доярок и пр. И. П. Павлов писал, что при первом знакомстве с новой средой неизбежно ожидать последствий всякого нового раздражения. Формируется оборонительное поведение, которое выражается пассивными и активными оборонительными реакциями, охраняющими животное от вредных факторов.
Пассивная оборонительная реакция у многих животных в первый момент выражается оцепенением, неподвижностью. Животные некоторых видов при сигнале опасности быстро разбегаются, прячутся и затаиваются. Оцепенение и затаивание связано с полным внешним торможением при доминировании пассивных оборонительных реакций (Л. Кру-шинский, 1943).
При активной оборонительной реакции животные изменяют позу. Например, быки копают ногами землю, ревут, выставляют вперед рога и бросаются на подходящего к ним человека или животного. Лошади прижимают уши к голове, оскаливают морду, храпят, взвизгивают, пытаются укусить, лягнуть задними ногами, а иногда ударить и топтать передними конечностями.
Большое значение в поведении животных имеют ориентировочные реакции. При вводе в новое помещение, появлении незнакомых людей у животных устанавливается характер стимулов, которые могут привести к проявлению, той или иной активности: оборонительной, пище- вой, половой и т. д. Они осматриваются, прислушиваются к голосам других животных, людей, обнюхивают землю, предметы, принюхиваются к доносящимся запахам, и в соответствии с данными ориентировки определяется то или иное поведение. Ориентировочные реакции особое место занимают в поведении молодых животных, у которых чаще, чем у взрослых, ориентировка сопровождается «любознательностью», рефлексом «что такое?». Исследовательская активность молодняка часто ведет к выработке новых навыков, которые затем осваиваются и взрослыми. Следовательно, элементы исследовательской активности прогрессивны и могут вести к совершенствованию поведения данной популяции животных.
Всем животным, особенно в молодом возрасте, свойственны игры — проявление игрового инстинкта. Проявление этого инстинкта важно для животных: давая выход избыточной энергии, игры служат тренировкой организма и развития тех форм поведения, которыми животное еще не обладает. Во время игры можно наблюдать и вспрыгивание молодых животных друг на друга. Это тоже является тренировкой к будущему маутингу — вспрыгиванию самца на самку во время спаривания. Строение тела копытных, их «жесткость» конструкции ограничивают разнообразие движений.
В связи с этим у лошадей, коров, овец, свиней и других копытных животных невозможно обхватывание тела самки (обнимательный рефлекс) и сразу происходит вспрыгивание (маутинг).
В формировании поведения особую роль играет подражание одного животного другому. Подражая движениям матери, других животных, молодое животное обучается кормиться, передвигаться, осматриваться и т. д. Подрастая, животные совершенствуют свои приемы добывания корма и другие необходимые для жизни действия.
Копытные животные, в том числе и сельскохозяйственные, в естественных условиях живут определенными группами. Поведение животных в больших группах определяется законом стадной иерархии, согласно которому каждое из них занимает свое ранговое место в группе. Такая организация помогала диким предкам животных выжить в борьбе за существование. Хотя условия существования современных домашних животных значительно изменились, все поведение скота, лошадей, свиней и овец при групповом содержании также определяется законами стада; всегда устанавливается иерархия — порядок подчинения нижестоящих особей вышестоящим по строго определенным ступеням (иерархическая лестница).
При формировании стад и табунов лидирующих животных сначала может быть несколько, но потом из них выделяется вожак, причем установление субординации происходит в ранговых боях. Ранг животного зависит от его силы, размеров, возраста.
Но во многих случаях, особенно у диких животных, лидером (вожаком) становится особь с большим житейским опытом. Знание закона стадной иерархии позволяет наиболее рационально формировать стада животных, производить перегруппировки.
Типы высшей нервной деятельности и поведение животных.
Тип нервной деятельности И. П. Павлов рассматривал как наследственную основу нервной деятельности — генотип, совокупность черт типа и изменений, обусловленных внешней средой,— фенотип. Животным каждого типа нервной деятельности свойственны различные черты поведения, обусловленные воспитанием в онтогенезе, условиями, в которых формируется деятельность организма.
Опыты по изолированному воспитанию щенков показали, что пассивные и активные оборонительные реакции, двигательная активность, злобность и другие показатели поведения могут сочетаться с различными типами нервной деятельности.
Различное воспитание не влияет на основные свойства нервных процессов: силу, уравновешенность и подвижность, но при изолированном воспитании чаще и интенсивнее проявляется пассивное оборонительное поведение, а активное оборонительное — реже.
Пассивное оборонительное поведение встречается у животных с сильными и слабыми нервными процессами, но слабость нервной системы служит фоном, более благоприятным для образования пассивного оборонительного поведения, которое усиливается изолированным воспитанием.
Свойства нервной системы не предопределяют никаких форм поведения, но составляют почву, на которой одни формы поведения складываются легче, другие — труднее.
Но в конфликтной ситуации и в экстремальных условиях животные разных типов неодинаково реагируют на обстановку, отвечая реакцией, свойственной качествам их нервной системы.
Применение учения И. П. Павлова и этологии в животноводстве. Условные рефлексы у животных приобретают практическое значение с первого дня жизни. Например, у поросят вырабатывают условные рефлексы на сосание одного определенного соска свиноматки. Это вызвано тем, что молочная продуктивность разных сосков у свиньи бывает неодинакова, поэтому для успешного выращивания поросят слабых целесообразно подсаживать к более молочным — передним, а более сильных — к задним соскам; таким образом вырабатывают условный рефлекс на сосание определенного соска. Этот рефлекс формируется очень быстро.
При помощи условных рефлексов можно облегчать процесс выращивания поросят-сосунов и отъемышей. Например, у поросят вырабатывают пищевой рефлекс на условный раздражитель — звук вентилятора. Для выработки и закрепления этого условного рефлекса достаточно одних суток. На звук вентилятора поросята встают, подбегают к дверце станка, поднимают визг, стремясь выйти и бежать к свиноматке. Кормление по сигналу проходит быстро и организованно. Опытные чабаны, учитывая способность коз быстро вырабатывать различные условные рефлексы, держат их в отарах овец в качестве вожаков. У коз вырабатывают условные рефлексы на специальные словесные (звуковые) команды, е соответствии с которыми козлы-вожаки изменяют направление своего движения при пастьбе, а овцы, обладающие близорукостью и пониженным слухом, но хорошо выраженным стадным инстинктом, следуют за ними. В результате вся отара движется в нужную сторону. Чабаны используют и другую поведенческую особенность овец, заключающуюся в том, что овцы ночью не покидают заслеженную территорию. Чтобы ночью овцы не разбрелись и не ушли, вечером отару несколько раз прогоняют по периметру пастбищного участка. В результате образуется заслеженная территория, за границы которой при ночной пастьбе овцы не заходят.
В коневодстве все производственные процессы: уход за лошадьми, тренинг и использование их в работе — построены на выработке многочисленных и многообразных условных рефлексов. Например, учитывая легкость образования двигательных условных рефлексов у лошадей на звуковые раздражители, табунщики для управления табуном на пастбище применяют словесные приказания и свистки. Выгон лошадей на пастбище и пригон их обратно сопровождаются свистком. Подгон уходящих из табуна лошадей связывается с другим сигналом. Условные рефлексы на различные звуковые сигналы вырабатываются очень быстро и облегчают труд табунщика.
studfiles.net
15. Внешние факторы инстинктивного поведения животных.
Внешние факторы инстинктивного поведения. Если задача внутренних факторов инстинктивного поведения состоит преимущественно в том, чтобы подготовить организм к выполнению определенного поведенческого акта, то внешние факторы чаще играют роль своеобразных активаторов этого инстинктивного действия.
Все инстинктивные действия заблокированы специальной системой, которая получила название «врожденный пусковой механизм». Это определенная совокупность нейросенсорных систем, обеспечивающих приуроченность поведенческих инстинктивных актов к ситуации, в которой такое поведение будет наиболее биологически адекватным, т. е. к так называемой «пусковой ситуации». Врожденный пусковой механизм реагирует на те или иные внешние раздражители или на их комбинации, для него характерна высокая избирательность. Каждый раздражитель, сигнал (или их комбинация) будут специфичны для определенной инстинктивной реакции. Врожденный пусковой механизм распознает их, проводит анализ, интегрирует информацию и снимает блокировку с данной реакции. При этом порог раздражимости соответствующих нервных центров снижается, и они активируются. Внутренняя мотивация «находит выход», и инстинктивная реакция осуществляется именно в тех условиях и в той ситуации, когда она биологически значима. Австрийский этолог К. Лоренц (1903–1989) назвал такой механизм «разблокирования» инстинктивной реакции врожденной схемой реагирования.
Инстинктивное действие проявляется в ответ на свой набор внешних раздражителей. Эти раздражители получили название «ключевые»,или «знаковые». Внешний сигнал в данном случае соотносят с ключом, который идеально подходит к замку (врожденному пусковому механизму). Например, в период размножения для самцов птиц ключевыми будут стимулы, характерные для самок того же вида, эти стимулы будут вызывать у самцов инстинктивные действия, связанные с ухаживанием, спариванием и т. д.
В качестве ключевых раздражителей могут выступать простые физические или химические признаки, их пространственные отношения (например, соотнесение размеров) или векторы.
Носителями ключевых раздражителей могут быть не только другие особи, но и растения, а также различные объекты неживой природы. Немецкий этолог Ф. Вальтер отметил, что у детенышей антилоп ключевым стимулом, определяющим выбор места отдыха, является любой вертикальный объект. Ключевой раздражитель выполняет здесь направляющую функцию.
Знаковые стимулы чрезвычайно разнообразны и по своей природе: они могут быть зрительными, акустическими, химическими и др. Например, в половом поведении многих насекомых, амфибий и некоторых млекопитающих ключевыми раздражителями служат химические вещества (половые аттрактанты, феромоны). К звуковым стимулам относятся разнообразные крики, песни, специфичные для определенного вида животных. Зрительные ключевые стимулы получили название «релизеры». К ним относятся различные морфологические признаки (особенности окраски тела, гребни, хохолки у птиц, наросты). Например, для самок крякв релизерами являются «зеркальца» на маховых перьях селезней. Существуют и особые видоспецифические комплексы движений, которые могут выступать в качестве знаковых стимулов (позы подчинения, угрожающие позы, приветственные ритуалы, брачные ритуалы).
Животное способно распознать ключевой раздражитель даже при первом предъявлении. Например, красное пятно на клюве чайки вызывает реакцию «попрошайничества» у птенцов. Для пояснения принципа действия этого раздражителя часто пользуются аналогией с ключом и замком.
Различают также настраивающие ключевые раздражители. Их действие отличается от знаковых раздражителей. Эти стимулы понижают порог раздражимости нервных центров и направляют ключевые раздражители.
Существование ключевых раздражителей и их роль в развитии инстинктивных реакций доказаны множеством наблюдений и опытов. Н. Тинберген изучал пищевую реакцию птенцов серебристых чаек и дроздов при появлении родительской особи методом макетов.
Естественная реакция голодного птенца чайки на родительскую особь заключается в том, что он клюет красное пятно на клюве (желтом) взрослой птицы. Тинберген в своих опытах использовал несколько макетов. Лишь один макет в точности повторял внешний облик головы взрослой серебристой чайки. На остальных макетах исключались отдельные детали, и постепенно макет становился все менее похожим на голову чайки. Последний макет представлял собой плоский красный предмет с продолговатым выступом. Однако реакция птенцов на этот предмет не только не была слабее реакции на первый макет, но даже превосходила ее. Еще более интенсивной становилась реакция птенца на макет в виде тонкой белой палочки с поперечными темно-красными полосами. Отсюда можно сделать вывод, что ключевыми раздражителями для появления реакции «попрошайничества» у птенцов серебристой чайки служат красный цвет и продолговатая форма.
В опытах с десятидневными птенцами дрозда в качестве макетов использовались плоские диски. Если птенцам дрозда предлагали круг, они тянулись к его верхней части, где предполагалась голова родительской птицы. Если к большому кругу прикрепляли маленький, птенцы начинали тянуться к нему, а когда прикрепляли два маленьких круга разного размера, решающей становилась относительная величина кругов. При большом размере «туловища» птенцы тянулись к дополнительному кругу больших размеров, при маленьком – к меньшему. Таким образом, ключевые раздражители в данном случае – взаимное расположение и относительная величина деталей макета.
Опыты по изучению ключевых стимулов у птиц проводили российские орнитологи Г.Л. Скребицкий и Т.И. Бибикова. В ходе опытов исследовалось отношение чайки к ее яйцам. Исследователи перекладывали яйца из одного гнезда в другое, заменяли их яйцами других видов птиц, иными предметами разной формы, величины, окраски. Чайки охотно начинали «высиживать» чужие яйца, а также яйца других птиц, различно окрашенные муляжи из разных материалов (стекло, глина и т. д.), посторонние предметы (шарики, картофелины, камни). Птицы не отказывались вкатывать в гнездо даже тяжелые каменные шарики, т. е. эту реакцию определял не вес «яйца». Г.Л. Скребицкий писал: «...чайки, сидящие на таких предметах, представляли очень оригинальную картину, но в особенности необычайным становилось зрелище, когда согнанная с гнезда птица возвращалась к нему обратно и, прежде чем сесть, заботливо поправляла клювом разноцветные шары, камешки или картошку».[8]
Птицы отказывались насиживать предметы, не имеющие округлой формы, например камни с острыми выступами или кубики. Ученые сделали вывод, что ключевыми стимулами для чайки были округлость предмета, отсутствие на нем выступов и углублений.
Если чайке предлагали два яйца разной величины, она начинала закатывать в гнездо более крупное. Исследователи наблюдали даже такую ситуацию, когда чайка пыталась насиживать деревянный макет яйца таких гигантских размеров, что она с трудом забиралась на него. В этой ситуации имеет место супероптимальная реакция. Животное сталкивается со сверхстимулом, который обладает супероптимальными признаками ключевого раздражителя, и начинает проявлять реакцию на него сильнее, чем в норме. Таким образом, ключевые раздражители подчинены закону суммации: с увеличением параметров раздражителя инстинктивная реакция увеличивается пропорционально. Это явление может служить объяснением повышенной реакции птенцов серебристой чайки на поперечно исчерченную красными полосками палочку.
На количественную сторону действия знаковых стимулов обратил внимание Н. Тинберген при изучении реакции преследования самки у самцов бабочки-бархатницы в период размножения. Наблюдения показали, что самец взлетает не только при приближении особей своего вида, но и при виде других летающих насекомых, а также небольших птиц и даже падающих с деревьев листьев. Ученый сделал вывод, что для бархатницы в этой ситуации первостепенное значение имеют какие-то зрительные ключевые раздражители. Химические стимулы в данном случае не могут быть знаковыми, потому что направление полета самцов никак не связано с направлением ветра, а значит, запахами они не руководствуются. Тинберген и его помощники изготовили макеты бабочек из бумаги и укрепили их на тонкой леске, привязанной к длинной удочке. У каждой серии макетов оставляли только один характерный внешний признак: цвет, размер, определенную форму. При подергивании удилища макетная бабочка начинала двигаться, что вызывало у самцов бархатницы ответную реакцию преследования. Интенсивность реакции фиксировалась наблюдением.
Результаты эксперимента показали, что реакцию преследования вызывали макеты всех цветов, но наиболее активно самцы следовали за черными «бабочками» – реакция на них была даже ярче выражена, чем при виде макетов коричневого цвета, который соответствовал естественной окраске самки бархатницы. В этом случае следует говорить об усилении зрительного стимула – темного цвета.
Аналогичная картина была получена при сравнении интенсивности реакции на размер макета. Самцы наиболее активно преследовали макеты большего размера, чем естественная величина самки. Такой стимул, как форма тела объекта, оказался для бархатниц не столь важен. Самцы реагировали на макеты всех форм, при этом наименее эффективны были длинные прямоугольные макеты. Однако наблюдения показали, что это связано с нарушением характера движения таких «бабочек»: он становился менее «танцующим».
Тинберген обратил внимание и на другую особенность действия ключевых стимулов, которую он назвал сумматором стимулирования. Ученый писал: «...слабо привлекающая белая модель вызовет тот же процент реакций, что и черная, если ее показывать на меньшем расстоянии, чем черную. Эффективность белой маленькой модели также заметно усиливается, если заставить ее „танцевать“. Таким образом, недостаточную эффективность одного параметра можно скомпенсировать усиленной привлекательностью совершенно иного параметра... стимулы складываются в некоем „сумматоре стимулирования“, который и заставляет бархатницу реагировать соответствующим образом».[9]
Кроме того, Тинберген отметил, что состояние самца определяет, какие стимулы в данный момент входят в этот сумматор. Например, в обычных условиях самцы реагировали только на тон окраски макета (темный или светлый), т. е. сами цвета в сумматор не входили. При кормлении же на макетах, окрашенных в яркий цвет, у самцов отмечалась реакция исключительно на голубые и желтые модели, т. е. знаковым стимулом становился цвет.
Реакция на ключевой стимул не всегда бывает адекватна ситуации и может не привести к желаемому результату. Так, Тинберген описывает явление, получившее название «осечки». Осечка в поведении животного происходит при столкновении со «сверхстимулом». Примером такого «сбоя» может служить выкармливание певчими птицами птенца кукушки. Ключевыми стимулами, заставляющими птицу-родителя кормить птенца, являются большой клюв и ярко окрашенный зев птенца. Оба эти признака у кукушонка имеют «сверхнормальное» выражение. Тинберген пишет: «Вполне возможно, что многие певчие птицы не только кормят птенца кукушонка, но и получают удовольствие из-за его огромного и привлекательного рта».[10]
Осечка может происходить и во взаимоотношениях представителей разных классов животных. Описан случай, когда птица кардинал на протяжении нескольких недель кормила насекомыми золотых рыбок в бассейне. Птица реагировала на широко открытый рот рыбок, являющийся для нее ключевым стимулом при выкармливании птенцов.
В заключение следует отметить, что инстинктивное поведение животных чаще всего обусловлено не отдельными факторами, а их комплексом. При этом необходимо сочетание внешних и внутренних факторов. Например, голуби кормят своих птенцов, отрыгивая «зобное молочко», богатое протеинами. Сам процесс образования в зобе «молочка» стимулируется выделением гормона пролактина (внутренний стимул). Однако реакция отрыгивания вызывается не наполнением зоба, а внешней стимуляцией со стороны птенца, который своей тяжестью давит на зоб родителя. В зимнее время даже при наполненном пищей зобе у голубя не происходит такой реакции, потому что нет внешней стимуляции.
studfiles.net